线粒体研究报告:急性 RyR1 Ca²⁺ 泄漏增强 NADH 依赖性线粒体呼吸能力
Nature communications:急性 RyR1 Ca²⁺ 泄漏增强 NADH 依赖性线粒体呼吸能力出处:Zanou N, Dridi H, Reiken S, et al. Acute RyR1 Ca²⁺ leak enhances NADH-linked mitochondrial respiratory capacity. Nat Commun. 2021; 12(1): 7219.
作者介绍:
持续性的兰尼碱受体Ca²⁺ 泄漏与心力衰竭或骨骼肌无力等病理状态密切相关。本研究发现,单次冲刺间歇训练(sprint interval training,SIT),而非中等强度持续训练(moderate-intensity continuous training,MICT),可在健康人体骨骼肌以及体外SIT模型(模拟 SIT 或 S-SIT)中诱导 RyR1 蛋白氧化和亚硝基化,进而导致 calstabin1解离。上述变化伴随着肌质网 Ca²⁺ 含量降低、线粒体氧化磷酸化相关蛋白水平升高、超复合体形成增加,以及 NADH 依赖性线粒体呼吸功能增强。在机制层面,(S-)SIT 可促进小鼠肌管和肌纤维中的线粒体 Ca²⁺ 摄取,并降低人类骨骼肌和小鼠肌管中丙酮酸脱氢酶的磷酸化水平。抑制 Ca²⁺ 泄漏或阻断线粒体 Ca²⁺ 摄取可削弱 S-SIT 诱导的适应性改变;这一结果亦得到蛋白质组学分析的支持。综上,本研究表明,在健康骨骼肌中诱导 RyR1 介导的急性、短暂性 Ca²⁺ 泄漏,可能有助于解释运动所产生的多重健康促进效应。兰尼碱受体(ryanodine receptors,RyRs)编码位于内质网/肌质网(endo/sarcoplasmic reticulum,ER/SR)上的细胞内 Ca²⁺ 释放通道¹。在哺乳动物中,RyRs 存在三种亚型。RyR1 和 RyR2分别参与骨骼肌和心肌的兴奋-收缩偶联²,并且也存在于非肌肉组织中³,而 RyR3 最初在脑组织中被鉴定出来⁴,随后发现其在多种组织中均有表达⁵。RyRs 是分子量超过 200 万道尔顿的同源四聚体,也是目前已知最大的离子通道。RyR1 和 RyR2 由两种 FKBP 编码蛋白稳定:calstabin1 和 calstabin2,即 FKBP12 和 FKBP12.6;其中,calstabin 意为钙通道稳定结合蛋白,这一名称反映了其在钙稳态中的作用⁶。RyRs 的功能还受到激酶⁷、磷酸酶⁸、磷酸二酯酶⁷˒⁹以及钙调蛋白⁷的进一步调控。RyR 的氧化、磷酸化和亚硝基化可导致 calstabin 从通道上解离,从而引发 Ca²⁺ 从ER/SR泄漏⁷。持续性的 Ca²⁺ 泄漏可使胞质和线粒体内 Ca²⁺ 水平过度升高,诱导活性氧(reactive oxygen species,ROS)生成增加、线粒体功能障碍以及细胞损伤¹⁰。该过程已被认为与心力衰竭¹¹、肌营养不良¹²、癌症相关肌无力¹³以及衰老¹⁴等病理状态有关。尽管持续性 RyR 泄漏可能参与多种病理过程,但近期在健康人群运动训练中的研究发现提示,运动过程中经肌肉内 RyR 通道发生的急性、短暂性 SR Ca²⁺ 泄漏,反而可能具有有益作用¹⁵,然而,这一观点仍有待进一步验证。运动在预防和治疗代谢性疾病方面的益处是毋庸置疑的¹⁶,但关于最佳运动类型组合,即为获得特定结果所需的运动量和运动强度,目前仍在讨论中。体育训练被定义为“在一定时间内定期重复进行的任何形式的身体运动”¹⁷。与 MICT 相比,高强度运动在改善超重或肥胖等代谢性疾病方面可能更为有效¹⁸˒¹⁹,这也是高强度间歇训练(high-intensity interval training,HIIT)目前越来越多地应用于患者护理中的原因²⁰–²²。SIT 是一种省时高效的运动方式,其可通过快速诱导骨骼肌代谢适应,使健康人群的最大有氧能力(VO₂peak)提高幅度与传统 MICT 相当²³˒²⁴。然而,触发这些有益效应的分子机制仍有待阐明,而 RyR1 功能/ Ca²⁺ 信号可能参与其中¹⁵˒²⁵˒²⁶。我们此前报道,单次 SIT 可改变人和小鼠肌纤维中的 RyR1 通道,并导致小鼠肌纤维静息胞质 Ca²⁺ 水平升高²⁶。此外,小鼠进行 3 周跑轮运动可诱导 RyR1 磷酸化,并与抗疲劳能力增强相关¹⁵。本文报道了一种分子机制:运动通过RyR1 诱导的急性、短暂性 SR Ca²⁺ 泄漏可引发骨骼肌线粒体重塑,从而改善线粒体呼吸功能。本文研究了进行单次 SIT 或 MICT 的人体受试者,并结合了来自人体骨骼肌活检、C2C12 肌管体外模拟 SIT 和 MICT(分别为 S-SIT 和 S-MICT),以及分离小鼠肌纤维离体 S-SIT 和 S-MICT 的数据。结果表明,单次(S-)SIT,而非(S-)MICT,可诱导RyR1 发生翻译后修饰(post-translational modifications,PTMs),导致calstabin1 从通道上解离,并降低 SR Ca²⁺ 含量。上述变化与线粒体 Ca²⁺ 摄取增加、丙酮酸脱氢酶(pyruvate dehydrogenase,PDH)编码蛋白 Ser293 位点磷酸化水平降低、线粒体氧化磷酸化(oxidative phosphorylation,OXPHOS)蛋白水平升高、线粒体超复合体(supercomplex,SC)形成增加,以及线粒体呼吸能力改善相关。通过使用 S107 改变 RyR1 Ca²⁺ 泄漏,并敲低线粒体 Ca²⁺ 单向转运体通道(mitochondrial Ca²⁺ uniporter channel,MCU)编码蛋白的实验结果表明,SIT 对线粒体适应性改变的积极作用,至少部分由特异性的 RyR1 Ca²⁺ 依赖性信号以及线粒体 Ca²⁺ 摄取介导;后者能够调控线粒体蛋白丰度及其功能。其中,S107 是一种 1,4-苯并硫氮䓬衍生物,被称为 Rycal,即一类可稳定 RyR1 关闭状态的新型药物,从而减少细胞内 Ca²⁺ 泄漏²⁷。
SIT 诱发人类骨骼肌中 RyR1 氧化和亚硝基化,以及 calstabin1 解离
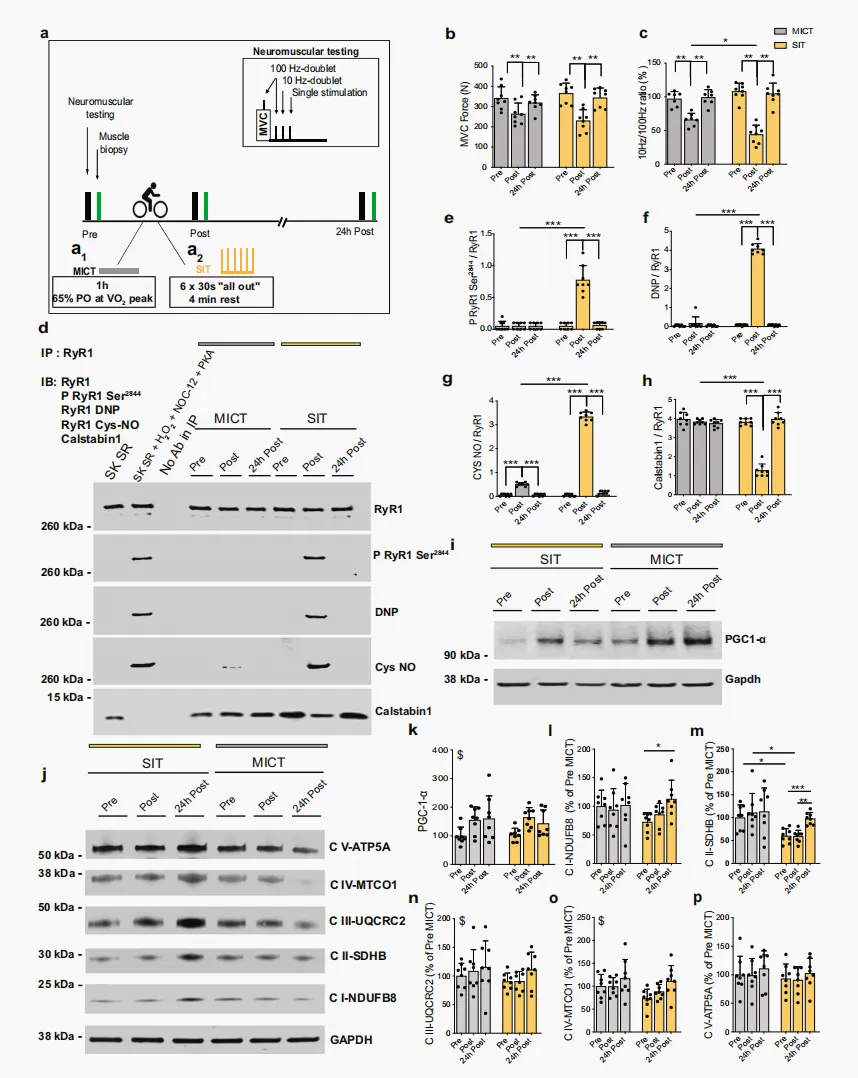
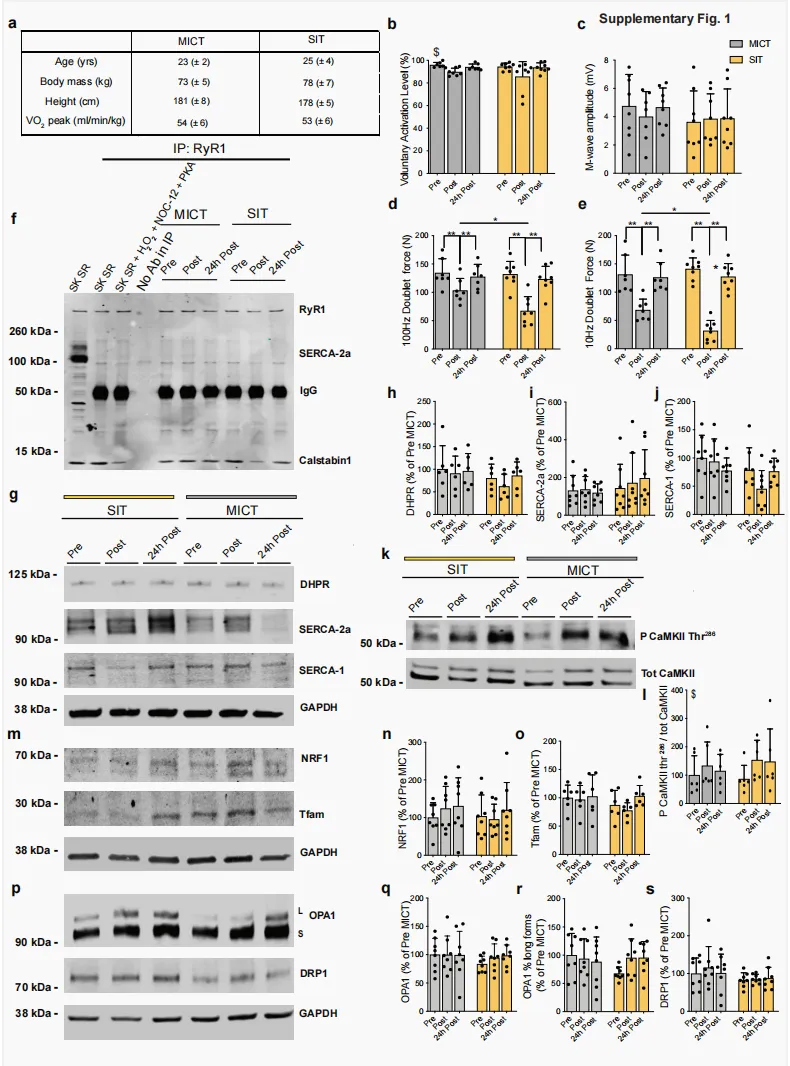
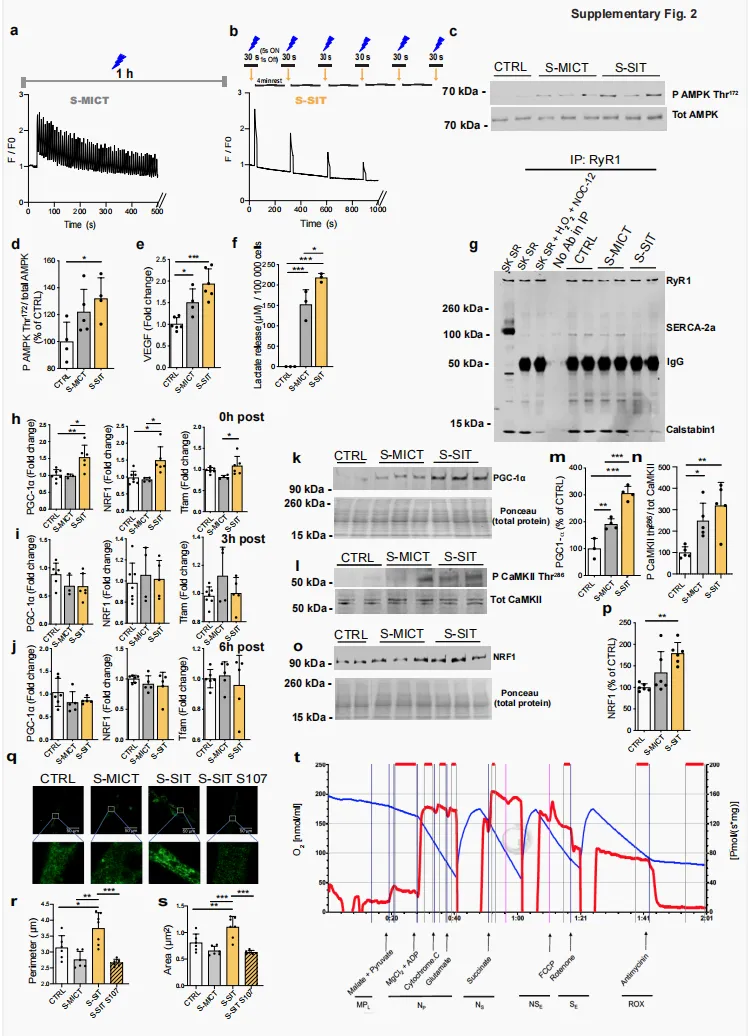
根据有氧适能水平匹配的休闲性运动志愿者(补充图 1a)完成单次 MICT 或 SIT。其中,MICT 为以 VO₂peak 对应功率输出的 65% 进行 1 h 自行车运动,总做功为680.9 ± 82.2 kJ(图 1a₁);SIT 为在功率自行车上完成 6 组 30 s “全力”骑行,每组间休息 4 min,6 组累计做功为 88.8 ± 18.0 kJ(图 1a₂)。在运动前(Pre)、运动后即刻(Post)以及运动后 24 h(24 h post),采用最大自主收缩(maximal voluntary contractions,MVC)结合电刺激的方法评估膝伸肌神经肌肉功能。分子层面的变化则通过采集股外侧肌活检样本进行研究,取样时间点分别为运动前(Pre)、运动后约 10 min(Post)以及运动后 24 h(24 h post)(图 1a)。与我们既往研究结果一致²⁶˒²⁸,单次 SIT 或 MICT 后即刻,MVC 力量下降约30%,并在运动后 24 h 完全恢复(图 1b)。我们对股神经进行电刺激,观察到膝伸肌最大自主激活水平存在小幅度下降,即轻度中枢性疲劳(补充图 1b),且该变化不依赖于运动强度。尽管未观察到 M 波振幅受损,即肌膜兴奋性得以维持(补充图 1c),但两组受试者在运动后即刻,100 Hz 和 10 Hz 电刺激诱发力均降低(补充图 1d、e),提示存在显著的外周性疲劳,即发生于肌纤维层面的疲劳。本研究还观察到,与 MICT 相比,SIT 后低频力下降更为明显,这体现在 10/100 Hz 比值降低(图 1c)。由于低频力下降与骨骼肌纤维 Ca²⁺ 处理受损有关²⁹,这提示,与 MICT 相比,SIT 可诱发更显著的 Ca²⁺ 稳态紊乱。随后,我们采用免疫共沉淀(immunoprecipitated,IP)实验,研究单次 SIT 或 MICT 是否会在人股外侧肌活检样本中诱发 RyR1 PTMs(包括磷酸化、亚硝基化或氧化)以及 calstabin1 解离;后者是 RyR 发生泄漏的标志性特征²⁷。SIT 后,肌肉活检样本显示 RyR1 氧化、亚硝基化和磷酸化显著增加,并伴随 calstabin1 解离(图 1d、e–h),提示 RyR1 可能处于泄漏状态。MICT 后,仅观察到 RyR1 轻度亚硝基化(图 1d、g)。尽管 MICT 诱导的 RyR1 亚硝基化在运动后即刻显著增加,但这一变化不足以诱导 calstabin1 从 RyR1 上解离(图 1d、g、h)。我们的 IP 样本中 RyR1 制备物具有特异性,因为肌/内质网Ca²⁺-ATP酶(sarco-endoplasmic Ca²⁺-ATPase,SERCA)蛋白仅在非 IP 样本中被检测到(补充图 1f)。随后,我们检测了二氢吡啶受体(dihydropyridine receptor,DHPR)和 SERCA 的蛋白水平,二者均参与肌肉兴奋-收缩偶联。SIT 和 MICT 均未显著改变 DHPR、SERCA2 或 SERCA1 的蛋白水平(补充图 1g–j)。这些结果表明,单次 SIT 而非 MICT 可在健康青年男性休闲运动者的骨骼肌中诱导 RyR1 氧化、亚硝基化和磷酸化,并伴随 calstabin1 解离,提示 RyR1 通道发生泄漏。图 1|单次 SIT 可诱导 RyR1 翻译后修饰,导致 calstabin1 从 RyR1 中解离,并提高人类骨骼肌中 OXPHOS 蛋白表达a 人体 MICT(a1)和 SIT(a2)模型。b,c b 膝伸肌 MVC 力评估;每组 n = 8;c 10 Hz 与 100 Hz 电刺激诱发力的比值;MICT 和 SIT 分别为 n = 7 和 8。采用双因素 ANOVA,并进行 Sidak 多重比较检验。d IP RyR1、RyR1 翻译后修饰及 calstabin1 解离的代表性 IB。DNP:RyR1 氧化。P RyR1 Ser2844:RyR1 Ser2844 位点磷酸化。Cys NO:RyR1 亚硝基化。SK SR:骨骼肌 SR 囊泡。SK SR经 200 µM H₂O₂、250 µM NOC-12 和 5 单位的 PKA 处理:RyR1 氧化、亚硝基化及 calstabin1 解离的阳性对照。IP 中无抗体:阴性对照 (d)。完整凝胶及额外对照见补充图 1f。e–h对(d)中 IB 的定量;每组 n = 8。采用双因素 ANOVA,并进行 Sidak 多重比较检验。i PGC-1α 的代表性 IB。j 线粒体 OXPHOS 蛋白的代表性 IB。所有 OXPHOS 蛋白裁剪部分均来自同一张印迹,见补充图5。k对(i)中 PGC-1α 蛋白进行定量,相对于 GAPDH 蛋白,并表示为 Pre-MICT 的百分比;每组 n = 8。采用双因素 ANOVA。l–p 对(j)中 OXPHOS 蛋白进行定量,相对于 GAPDH 蛋白,并表示为 Pre-MICT的百分比;每组 n = 8。采用双因素 ANOVA,并进行 Sidak 多重比较检验(l,m)。数据以 mean ± SD 表示。*p ≤ 0.05,**p ≤ 0.01,***p ≤ 0.001,$表示时间主效应。源数据作为 Source Data 文件提供。补充图 1|参与者特征、膝伸肌神经肌肉功能,以及人类骨骼肌中的 SR 和线粒体蛋白a 参与研究志愿者的生理特征;每组 n = 8。数值为 mean ± SD。b–e 膝伸肌最大自主激活水平(b)、M 波幅度(c)以及股神经分别接受 100 Hz 和 10 Hz 成对刺激时的诱发力(分别为 d 和 e);每组 n = 8。采用双因素 ANOVA,并进行 Sidak 多重比较检验(d 和 e)。f 非 IP 和 IP RyR1 样本的IB,以及 RyR1、SERCA2a 和 calstabin1 的 IB。显示 IgG 用于区分 IP 样本和 SERCA2a 阳性对照(第一泳道)。SK SR:骨骼肌 SR 囊泡。SK SR 经 200 µM H₂O₂、250 µM NOC-12 和 5 单位 PKA 处理:calstabin1 解离的阳性对照。IP 中无抗体:RyR1 IP 的阴性对照。g–j DHPR(g,h)、SERCA2a(g,i)和 SERCA1(g,j)蛋白的代表性 IB 及定量,相对于 GAPDH 蛋白;每组 n = 8。采用双因素 ANOVA,并进行 Sidak 多重比较检验(j)。k,l Thr286 位点磷酸化 CaMKII(相对于总蛋白)与总 CaMKII(相对于总蛋白)比值的代表性 IB(k)及定量(l);每组 n = 6。采用双因素ANOVA。m–o NRF1(m,n:每组 n = 8)和 Tfam(m,o:每组 n = 6)蛋白的代表性 IB 及定量,相对于 GAPDH 蛋白。采用双因素 ANOVA。p–s OPA1 长、短形式(q,r:每组 n = 8)和 DRP1(s:每组 n = 8)的代表性 IB(p)及定量,相对于 GAPDH 蛋白。采用双因素 ANOVA。数据以 mean ± SD 表示。*p ≤ 0.05,**p ≤ 0.01,$表示时间主效应。源数据作为 Source Data 文件提供。
SIT 诱导 RyR1 修饰后,人类肌肉活检中线粒体 OXPHOS 蛋白水平升高
运动训练可提高人类骨骼肌线粒体呼吸能力³⁰˒³¹。这一结果来源于线粒体体积增加、OXPHOS 蛋白水平升高,以及线粒体电子传递系统(electron transfer system,ETS)功能和效率的改善³²。过氧化物酶体增殖物激活受体γ共激活因子 1α(peroxisome proliferator-activated receptor co-activator 1α,PGC-1α)在激活线粒体生物发生所需基因方面具有重要作用,并与核呼吸因子(nuclear respiratory factor,NRF)和线粒体转录因子 A(mitochondrial transcription factor A,Tfam)协同发挥作用³³˒³⁴。耐力运动可在人类骨骼肌中诱导 PGC-1α 信使 RNA(messenger RNA,mRNA)出现显著而短暂的升高³⁵,即使单次耐力运动也可增加人类骨骼肌中 PGC-1α mRNA 和蛋白水平³⁶。我们使用一种可识别所有 PGC-1α 亚型的抗体³⁷进行检测,结果观察到,单次 MICT 和 SIT 均可在运动后诱导 PGC-1α 蛋白水平升高(图 1i、k)。
肌肉收缩过程中,由于 Ca²⁺ 从 SR 释放,胞质 Ca²⁺ 水平显著升高。值得注意的是,已有研究表明,Ca²⁺/钙调蛋白依赖性蛋白激酶II(Ca²⁺/calmodulin-dependent protein kinase II,CaMKII)可响应胞质 Ca²⁺ 水平升高而触发 PGC-1α 表达³⁸。SIT 和 MICT 均可诱导 CaMKII 磷酸化水平相似程度地升高(补充图 1k、l)。NRF1 和 Tfam 蛋白水平未观察到显著变化(补充图 1m–o)。
随后,我们研究了 SIT 肌肉中的 RyR1 修饰是否与 OXPHOS 蛋白水平升高相关。单次 SIT,而非 MICT,可显著增加线粒体 OXPHOS 复合体I(complex I,CI)和复合体II(complex II,CII)水平,而其他复合体在两组之间未见差异(图 1j、l–p)。运动应答过程中,线粒体还可通过分裂和融合发生动态重塑³⁹。线粒体内膜融合过程由视神经萎缩蛋白1(optic atrophy 1,OPA1)介导。OPA1 具有 5 种形式,其中 2 种长形式促进融合,3 种短形式促进分裂⁴⁰;而线粒体外膜融合则由线粒体融合蛋白1(mitofusin 1,MFN1)和线粒体融合蛋白2(mitofusin 2,MFN2)介导⁴¹。GTP酶动力蛋白相关蛋白1(dynamin-related protein 1,DRP1)则介导线粒体分裂⁴⁰。已有研究表明,高强度、大运动量训练后 3 h,OPA1 总蛋白水平升高,但 SIT 后 3 h 未出现这一变化⁴²。同时,SIT 对 OPA1 各种形式的影响尚未被报道。在本研究中,我们观察到,单次 SIT 或 MICT 后,OPA1 总蛋白及其不同形式均未发生变化(补充图 1p–r)。SIT 或 MICT 后,DRP1 蛋白水平亦未发生改变(补充图 1p、s)。
S-SIT 诱导 RyR1 修饰和 calstabin1 解离,并伴随 SR 中 Ca²⁺ 含量下降
在人类骨骼肌活检样本中,我们发现,单次 SIT 而非 MICT 可诱导 RyR1 蛋白修饰和 calstabin1 解离,并伴随线粒体 OXPHOS 水平升高。由于人体样本的获取和可用数量有限,某些技术难以应用于人体样本,因此,为进一步探究这些 RyR1 修饰与线粒体适应性变化之间的潜在联系,我们建立了 SIT 和 MICT 的细胞模型,即 S-SIT 和 S-MICT。我们分别基于高频和低频刺激模式,对小鼠 C2C12 肌管进行特异性刺激⁴³,以期诱导出与人体样本中观察到的相似变化。之所以选择小鼠 C2C12 肌管,是因为其是体外模拟运动的有效模型⁴⁴。从代谢角度来看,C2C12 肌管是目前最接近成人骨骼肌的可用细胞模型。与大鼠骨骼肌细胞培养物或人原代骨骼肌细胞培养物相比,C2C12 肌管与人类骨骼肌具有更多共同特征,因此更适用于运动相关研究⁴⁵。此外,我们还采用了从小鼠趾短屈肌(flexor digitorum brevis,FDB)中分离获得的离体单根肌纤维作为补充模型。选择这些模型的原因在于,SIT,即重复性“全力”冲刺运动,无法在小鼠体内实现。我们未采用来源于人原代成肌细胞的肌管,因为这类肌管不能模拟成人骨骼肌的 Ca²⁺ 处理过程,并且缺乏收缩能力⁴⁶。为探究 RyR1 泄漏状态是否与所观察到的线粒体 OXPHOS 能力升高相关,我们首先采用电刺激在 C2C12 肌管中诱导 S-MICT 和 S-SIT 收缩(图 2a)。正如刺激模式所预期的那样,S-SIT 和 S-MICT 诱导的 Ca²⁺ 释放模式不同(分别见补充图 2a、b)。运动诱导代谢应激的一个关键标志是 AMP 激活蛋白激酶(AMP-activated protein kinase,AMPK)磷酸化⁴⁷。在我们的细胞模型中,S-SIT 显著增加了 AMPK Thr172 位点的磷酸化水平(补充图 2c、d),这与在人类骨骼肌中的报道一致³¹。对刺激后即刻收集的全细胞裂解液进行代谢组学分析显示,S-SIT 中 AMP/ATP 和 ADP/ATP 比值明显升高,这支持了 S-SIT 肌管中 AMPK 磷酸化水平升高的结果(图 2b)。磷酸肌酸/肌酸比值变化不大,且未观察到 NAD 或 NADH 水平发生变化(图 2b)。与对照组相比,两种模型均表现出血管内皮生长因子 mRNA 水平显著升高(补充图 2e)以及乳酸释放增加(补充图 2f),且 S-SIT 的效应强于 S-MICT。综上,上述结果验证了我们所建立的体外模型能够模拟这两种运动模式,并可用于后续机制研究。随后,我们研究了 S-SIT 和 S-MICT 诱导下 RyR1 PTMs 以及 RyR1 与 calstabin1 结合状态的变化。与在人类骨骼肌活检样本中观察到的结果一致,单次 S-SIT 而非 S-MICT 可诱导 RyR1 氧化和亚硝基化,并导致 calstabin1 解离(图 2c–f),提示C2C12 肌管在 S-SIT 刺激后同样出现 RyR1 泄漏状态。与人体数据类似,在细胞模型中获得的 RyR1 免疫沉淀制备物未受到 SERCA 污染(补充图 2g)。随后,我们在这些体外模型中进一步研究 SR Ca²⁺ 储存是否发生改变。首先,我们使用 RyR1 激动剂咖啡因,检测 S-SIT 或 S-MICT 后肌管中 SR 可释放的最大 Ca²⁺ 含量。电刺激后即刻,在无细胞外 Ca²⁺ 条件下进行咖啡因刺激,即避免可能由质膜通道改变所带来的混杂影响,结果显示,与对照组和 S-MICT 肌管相比,S-SIT 肌管 SR 释放的 Ca²⁺ 更少,提示其 SR Ca²⁺ 含量降低(图 2g、h)。Ca²⁺ 可在 SR 内通过与无机磷酸盐结合而被螯合,而无机磷酸盐会在运动过程中积累⁴⁸;此外,Ca²⁺ 也可被肌钙集蛋白螯合⁴⁹,从而限制 Ca²⁺ 从 SR 释放。由于这些机制可能影响上述测量结果,我们进一步直接检测了 S-SIT 和 S-MICT 后的 SR Ca²⁺ 含量。我们将一种靶向肌质网的比率型 cameleon 探针(D1ER)质粒转染至分化后的肌管中,以直接评估 SR Ca²⁺ 水平。S-SIT 和 S-MICT 后即刻,采用荧光共振能量转移(fluorescence resonance energy transfer,FRET)系统对静息状态下的 SR Ca²⁺ 储备进行定量⁵⁰。结果显示,与 S-MICT 和对照肌管相比,S-SIT 肌管中反映 SR Ca²⁺ 水平的 D1ER 信号降低(图 2i、j),表明单次 S-SIT 后,RyR1 泄漏导致 SR Ca²⁺ 含量下降。为排除 S-SIT 和 S-MICT 肌管中钙库操控性钙内流(store-operated Ca²⁺ entry,SOCE)的任何变化对 SR Ca²⁺ 泄漏的影响,我们检测了 STIM1 水平以及 SOCE Ca²⁺ 瞬变。其中,STIM1 是骨骼肌 SOCE 的关键调控因子⁵¹。与对照组相比,两种条件下 STIM1 表达和 SOCE 均未发生变化(图 2k–o)。总体而言,我们在 C2C12 肌管中的结果表明,S-SIT 可诱导与人体 SIT 后骨骼肌中相似的 RyR1 修饰,并提示单次 S-SIT 后肌管 SR Ca²⁺ 含量降低;这一变化很可能是 calstabin1 从 RyR1 解离所引发 Ca²⁺ 泄漏的结果。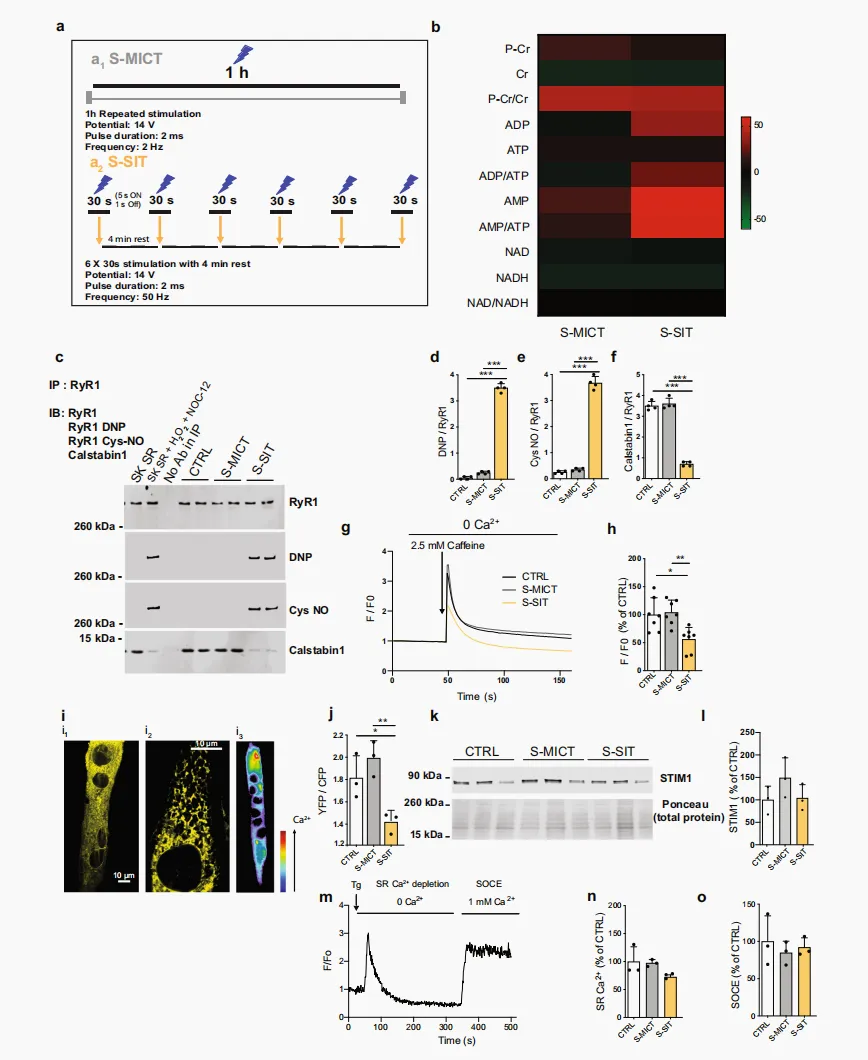 图 2|S-SIT 在 C2C12 肌管中诱导 RyR1 翻译后修饰、calstabin1 解离,并降低 SR Ca²⁺ 含量a C2C12 肌管中 S-MICT(a1)和 S-SIT(a2)模型。b 刺激后立即进行的代谢组学分析,变化以CTRL 百分比表示;每组 n = 2 独立生物学实验,每个实验 6 个重复。c–f RyR1 免疫沉淀的代表性 IB(c)及定量分析(d–f),评估刺激后 RyR1 翻译后修饰。DNP:RyR1 氧化。Cys NO:RyR1 硝基化。SK SR:骨骼肌 SR 囊泡。SK SR 经 200 µM H₂O₂ 和 250 µM NOC-12 处理:RyR1 氧化、硝基化及 calstabin1 解离的阳性对照。IP 中无抗体:阴性对照;每组 n = 4 独立生物学实验。采用单因素ANOVA 并进行 Tukey 多重比较检验,以及 Kruskal–Wallis ANOVA 后 Dunn 多重比较检验(e)。完整凝胶及额外对照见补充图 2g。g–h 刺激后立即响应 2.5 mM 咖啡因的 Fluo-4 荧光原始记录(g)及定量(h);每组 n = 7 独立生物学实验。单因素 ANOVA 并进行 Tukey 多重比较检验。i,j 基于荧光共振能量的 SR Ca²⁺ 含量评估(i)及 D1ER 比值定量(j),刺激后立即进行 S-MICT 和 S-SIT 实验。(i1)535 nm 下 D1ER 质粒分布记录。(i2)D1ER 信号网状模式细节。(i3)Ca²⁺ 分布伪彩色显示(从深蓝低 Ca²⁺ 到红色高 Ca²⁺ ,见箭头);每组 n = 3 独立生物学实验。单因素 ANOVA 并进行Tukey 多重比较检验。k–l 刺激后 S-MICT 和 S-SIT 肌管中 STIM1 的代表性 IB(k)及定量(l);每组 n = 3 独立生物学实验。单因素 ANOVA。m–o 刺激后 S-MICT 和 S-SIT 肌管的 SOCE 表征。SOCE 原始记录(m);SR Ca²⁺ 库定量(n)及 SOCE 定量(o);每组 n = 3 独立生物学实验。单因素 ANOVA。数据以 mean ± SD 表示。*p ≤ 0.05,**p ≤ 0.01,***p ≤ 0.001。源数据作为 Source Data 文件提供。补充图 2|体外 S-MICT 和 S-SIT 模型的特征
图 2|S-SIT 在 C2C12 肌管中诱导 RyR1 翻译后修饰、calstabin1 解离,并降低 SR Ca²⁺ 含量a C2C12 肌管中 S-MICT(a1)和 S-SIT(a2)模型。b 刺激后立即进行的代谢组学分析,变化以CTRL 百分比表示;每组 n = 2 独立生物学实验,每个实验 6 个重复。c–f RyR1 免疫沉淀的代表性 IB(c)及定量分析(d–f),评估刺激后 RyR1 翻译后修饰。DNP:RyR1 氧化。Cys NO:RyR1 硝基化。SK SR:骨骼肌 SR 囊泡。SK SR 经 200 µM H₂O₂ 和 250 µM NOC-12 处理:RyR1 氧化、硝基化及 calstabin1 解离的阳性对照。IP 中无抗体:阴性对照;每组 n = 4 独立生物学实验。采用单因素ANOVA 并进行 Tukey 多重比较检验,以及 Kruskal–Wallis ANOVA 后 Dunn 多重比较检验(e)。完整凝胶及额外对照见补充图 2g。g–h 刺激后立即响应 2.5 mM 咖啡因的 Fluo-4 荧光原始记录(g)及定量(h);每组 n = 7 独立生物学实验。单因素 ANOVA 并进行 Tukey 多重比较检验。i,j 基于荧光共振能量的 SR Ca²⁺ 含量评估(i)及 D1ER 比值定量(j),刺激后立即进行 S-MICT 和 S-SIT 实验。(i1)535 nm 下 D1ER 质粒分布记录。(i2)D1ER 信号网状模式细节。(i3)Ca²⁺ 分布伪彩色显示(从深蓝低 Ca²⁺ 到红色高 Ca²⁺ ,见箭头);每组 n = 3 独立生物学实验。单因素 ANOVA 并进行Tukey 多重比较检验。k–l 刺激后 S-MICT 和 S-SIT 肌管中 STIM1 的代表性 IB(k)及定量(l);每组 n = 3 独立生物学实验。单因素 ANOVA。m–o 刺激后 S-MICT 和 S-SIT 肌管的 SOCE 表征。SOCE 原始记录(m);SR Ca²⁺ 库定量(n)及 SOCE 定量(o);每组 n = 3 独立生物学实验。单因素 ANOVA。数据以 mean ± SD 表示。*p ≤ 0.05,**p ≤ 0.01,***p ≤ 0.001。源数据作为 Source Data 文件提供。补充图 2|体外 S-MICT 和 S-SIT 模型的特征a,b 响应 S-MICT(a)和 S-SIT(b)刺激的标准化 Fluo-4 胞质荧光。c,d 刺激后即刻 Thr172 位点磷酸化 AMPK 和总 AMPK 的代表性 IB(c)及定量(d);每组 n = 4 独立生物学实验。采用单因素ANOVA,并进行 Tukey 多重比较检验。e VEGF 相对于 Cyclophilin B 的 mRNA 定量;n = 6(CTRL、S-SIT)和 4(S-MICT)个独立生物学实验。采用单因素 ANOVA,并进行 Tukey 多重比较检验。f 刺激后即刻乳酸释放量定量;每组 n = 3 独立生物学实验。采用单因素 ANOVA,并进行 Tukey 多重比较检验。g 非 IP(第一泳道)和 IP RyR1 样本的 IB,以及 RyR1、SERCA2a 和 calstabin1 的 IB。显示 IgG 用于区分 IP 样本和 SERCA2a 阳性对照(第一泳道)。SK SR:骨骼肌 SR 囊泡。SK SR 经 200 µM H₂O₂、250 µM NOC-12 处理:calstabin1 解离的阳性对照。IP 中无抗体:RyR1 IP 的阴性对照。h–j 刺激后即刻(h:n = 7 CTRL、6 S-SIT、3 S-MICT 独立生物学实验)、3 h 和 6 h(i, j:n = 7 CTRL、6 S-SIT、4 S-MICT 独立生物学实验)PGC-1a、NRF1 和Tfam 相对于 Cyclophilin B 的 mRNA 水平。采用单因素 ANOVA,并进行 Tukey 多重比较检验(h)。k–m 刺激后 72 h PGC-1a 蛋白相对于总蛋白的 IB(k)及定量(m);n = 3(CTRL)和 4(S-MICT、S-SIT)个独立生物学实验。采用单因素 ANOVA,并进行 Tukey 多重比较检验。l–n Thr286 位点磷酸化 CaMKII 和总 CaMKII 的代表性 IB(l)及定量(n)(二者均相对于总蛋白);每组 n = 5 独立生物学实验。采用单因素 ANOVA,并进行 Tukey 多重比较检验。o,p 刺激后 72 h NRF1 蛋白相对于总蛋白的 IB(o)及定量(p);每组 n = 5 独立生物学实验。采用单因素 ANOVA,并进行 Tukey 多重比较检验。q 刺激后 72 h 未使用或使用 10 µM S107 处理时的 Mitotracker green荧光。r,s 线粒体周长和面积的定量;n = 6(CTRL、S-MICT、S-SIT S107)和 7(S-SIT)个独立生物学实验。采用单因素 ANOVA,并进行 Tukey 多重比较检验。t 肌管线粒体呼吸能力的原始记录。数据以 mean ± SD 表示。*p ≤ 0.05,**p ≤ 0.01,***p ≤ 0.001。源数据作为 Source Data 文件提供。
RyR1 泄漏状态与线粒体 OXPHOS 蛋白表达、SC 形成和 NADH 依赖性呼吸能力的增加有关
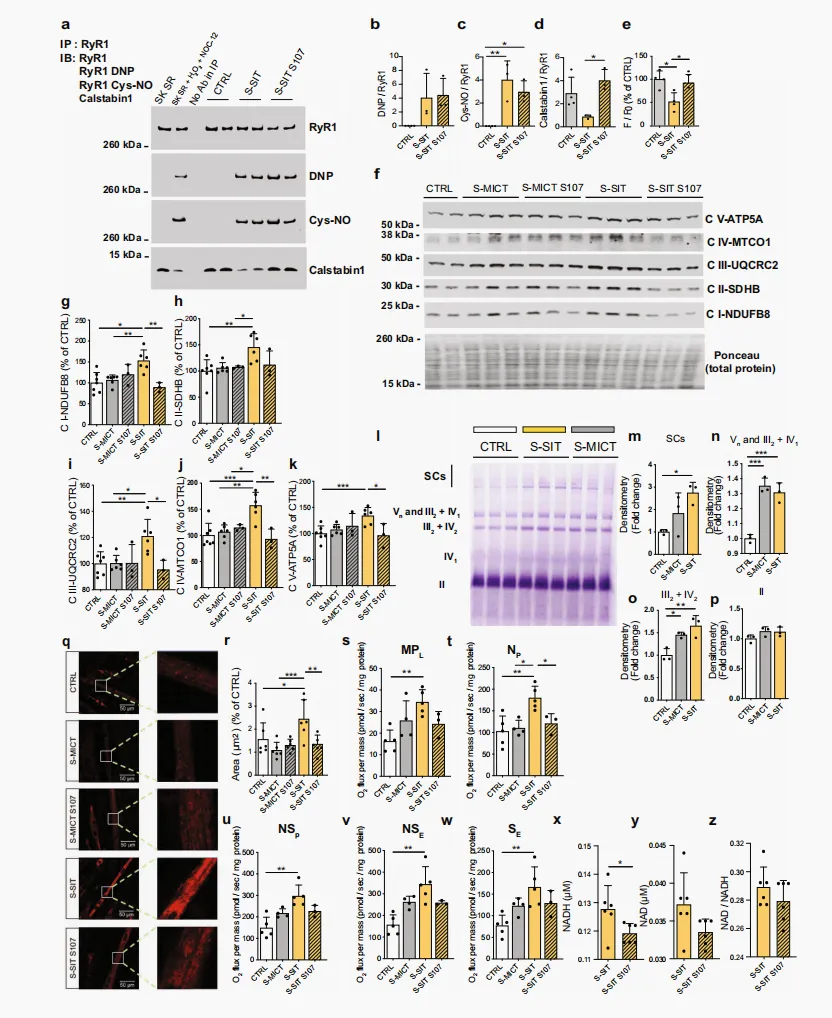
为研究 RyR1 Ca²⁺ 泄漏与线粒体适应之间的联系,我们在 S-SIT 模型中使用 S107⁵² 降低 RyR1 Ca²⁺ 泄漏,并评估线粒体蛋白的反应。S107 通过增强 RyR1 与 calstabin1 之间的物理相互作用发挥作用,从而预防/阻止 Ca²⁺ 泄漏⁵³。我们发现,在我们的 S-SIT 肌管中,10 μM 的 S107 处理足以使 calstabin1 重新结合至 RyR1。S107 处理 3 h 后恢复了 calstabin1 与 RyR1 的结合,但不影响 RyR1 的 PTMs(图 3a–d),这与既往研究一致¹¹˒⁵²。S107 处理还逆转了 S-SIT 肌管细胞中 SR Ca²⁺ 储备下降的现象(图 3e)。在线粒体 OXPHOS 蛋白方面,其在刺激后 72 h 的 S-SIT 肌管中显著增加,但在S-MICT 肌管中未见增加;而 S107 处理(刺激后立即开始并持续 72 h)削弱了 S-SIT 肌管中的这些适应性变化(图 3f–k)。ETS 可形成称为 SCs 的超复合体结构⁵⁴,而这些结构可直接提高代谢能力和效率⁵⁵。运动训练会诱导人体骨骼肌中 SCs 的形成,作为满足能量需求增加的一种适应性机制⁵⁶。我们研究了单次刺激后 72 h 时,S-SIT 和 S-MICT 肌管中 SCs 的形成情况。S-SIT 和 S-MICT 均增加了总体 SC 含量以及特定线粒体复合物的形成(图 3l)。虽然 S-SIT 和 S-MICT 均诱导了复合物 Vn + III₂ + IV₁ 和 III₂ + IV₂ 的形成,且游离复合物 II 未发生变化(图 3n–p),但蓝色非变性聚丙烯酰胺凝胶电泳(blue native polyacrylamide gel electrophoresis,BN-PAGE)定量结果显示,在 C2C12 细胞中,S-SIT 在组装主要由复合物 I + III + IV 构成的 SCs 方面更为有效(图 3l、m)⁵⁷。与人体骨骼肌中的研究类似,我们也在受刺激的肌管中检测了线粒体生物发生标志物。S-SIT 后即刻,肌管中 PGC-1α、NRF1 和 Tfam 的 mRNA 水平均显著升高,而 S-MICT 后未见显著升高(补充图 2h–j)。使用与人体样本相同的抗体,我们观察到,在 S-SIT 后 72 h,S-SIT 和 S-MICT 肌管中的 PGC-1α 均显著增加,但 S-SIT 肌管中的增加幅度更大(补充图 2k、m)。两种细胞中 CaMKII 磷酸化水平相近且均升高,也支持了这一结果(补充图 2l、n)。NRF1 蛋白仅在 S-SIT 肌管中显著增加(补充图 2o、p)。使用 Mitotracker red 进行线粒体染色显示,与 S-MICT 和对照肌管相比,S-SIT 肌管中的线粒体面积显著增加(图 3q、r)。已知 Mitotracker red 依赖于线粒体膜电位。因此,为排除线粒体膜电位可能对线粒体面积测定造成的混杂影响,我们还使用了Mitotracker green 探针,结果进一步证实了上述发现(补充图 2q–s)。重要的是,S107 处理削弱了 S-SIT 肌管中的这些变化(图 3q、r 及补充图 2q–s)。线粒体相关基因/蛋白表达的变化以及酶活性测定,只能为线粒体含量和氧化能力提供静态的替代性指标,并不能反映线粒体功能的复杂性⁵⁸。目前,关于间歇性高强度训练后线粒体蛋白含量及功能变化的研究仍较少³²。我们采用高分辨率呼吸测定技术,探究上述发现是否伴随着线粒体呼吸功能的改善(补充图 2t)⁵⁹。在非磷酸化状态下,使用苹果酸和丙酮酸评估的泄漏状态(MPL)显示,与 S-MICT 和对照肌管相比,S-SIT 组显著升高(图 3s)。通过加入饱和浓度的二磷酸腺苷(adenosine diphosphate, ADP),随后加入谷氨酸测定的OXPHOS 状态(N-依赖性通路(NP))显示,与对照肌管相比,S-SIT 组显著升高,而 S-MICT 组未观察到明显变化(图 3t)。进一步加入琥珀酸以诱导电子汇入 Q 连接点,从而增加耗氧量(NADH 和琥珀酸依赖性通路,NSP 状态),S-SIT 肌管的通量高于 S-MICT 和对照肌管(图 3u)。当加入羰基氰化物-4-三氟甲氧基苯腙(carbonyl cyanide 4-(trifluoromethoxy)phenylhydrazone,FCCP)以评估最大 ETS 活性(NSE 状态)时,仅 S-SIT 肌管的最大呼吸能力显著升高(图 3v)。使用鱼藤酮抑制 NADH 依赖性呼吸后,进一步突出了非磷酸化琥珀酸依赖性通路(SE)的增加(图 3w)。重要的是,S107 处理特异性削弱了 S-SIT 肌管中的 NADH 依赖性呼吸通路(图 3t)。这些结果表明,单次 S-SIT 可增加线粒体蛋白,并增强线粒体呼吸功能。S-SIT S107 肌管在刺激后 72 h 时,与 S-SIT 肌管相比,总 NADH 水平出现显著下降(图 3x),而 NAD 和 NAD/NADH 水平相近(图 3y、z)。尽管线粒体区室内的 NADH/NAD⁺ 比值高于胞质,但在本研究条件下观察到的 NADH 水平变化应被视为全局性改变,而非线粒体特异性改变。综合来看,我们的结果提示,S-SIT 与 S-SIT S107 肌管在线粒体 NADH 相关底物驱动的呼吸速率方面存在特异性差异。S-SIT 处理诱导了总体 SCs,即由复合物 I + III + IV 构成的超复合物的增加,这进一步支持了上述结果(图 3l、m)。总体而言,我们的结果表明,S-SIT 诱导的线粒体向有氧代谢能力增强表型的有益适应,至少部分由 RyR1/calstabin1 解离引起的急性 SR Ca²⁺ 泄漏所驱动。
图 3|相比 MICT,S-SIT 在 C2C12 肌管中能诱导更高的线粒体蛋白含量、超复合体水平和呼吸能力,这些效应可被 S107 诱导的 RyR1 稳定所抑制a–d 刺激后 3 h 未处理或 S107 处理 3 h 的 IP RyR1 翻译后修饰代表性 IB(a)及定量分析(b–d);n = 4(CTRL)和 3(S-MICT、S-SIT)独立生物学实验。单因素 ANOVA,并进行 Tukey 多重比较检验(c,d)。e 刺激后 3 h 未处理或 S107 处理的 Fluo-4/AM 荧光比值定量;n = 4(CTRL、S-SIT)和 3(S-SIT S107)独立生物学实验。单因素 ANOVA 并进行 Tukey 多重比较检验。f–k 刺激后 72 h 未处理或 S107 处理 72 h 的线粒体 OXPHOS 蛋白代表性 IB(f)及定量分析(g–k)。完整OXPHOS 印迹见补充图6;n = 7(CTRL)、6(S-MICT、S-SIT)和 3(S-MICT S107、S-SIT S107)独立生物学实验。单因素 ANOVA 并进行 Tukey 多重比较检验。l–p 刺激后的超复合体 IB(l)及定量分析(m–p);每组 n = 3 独立生物学实验。单因素 ANOVA 并进行 Tukey 多重比较检验(m–o)。q,r 刺激后 72 h Mitotracker red 荧光(q)及线粒体面积定量(r),未处理或 S107 处理;每组 n = 6 独立生物学实验。单因素 ANOVA 并进行 Sidak 多重比较检验。s–w 刺激后 72 h 及 S107 处理的单位蛋白 O₂ 通量(pmol/s/mg);s MPL:苹果酸-丙酮酸泄漏状态;t NP:ADP 刺激的 N-OXPHOS 状态;u NSP:N 与 S-OXPHOS 途径;v NSE:ET 状态,非偶联;w SE:S 途径;n = 5(CTRL、S-SIT)、4(S-MICT)和 3(S-SIT S107)独立生物学实验。单因素 ANOVA 并进行 Sidak 多重比较检验。x–z 刺激后 72 h S-SIT 肌管中未处理或 S107 处理的 NADH(x)、NAD(y)浓度及 NADH/NAD 比值(z);每组 n = 6 独立生物学实验。未配对 t 检验。S107 组在刺激后立即给予 10 µM S107 处理至指定时间。数据以 mean ± SD 表示。*p ≤ 0.05,**p ≤ 0.01,***p ≤ 0.001。源数据作为 Source Data 文件提供。 图 6|C2C12 细胞 OXPHOS 蛋白的原始凝胶图
图 6|C2C12 细胞 OXPHOS 蛋白的原始凝胶图 减少 RyR1 Ca²⁺ 泄漏会抑制 S-SIT 的其他生理适应
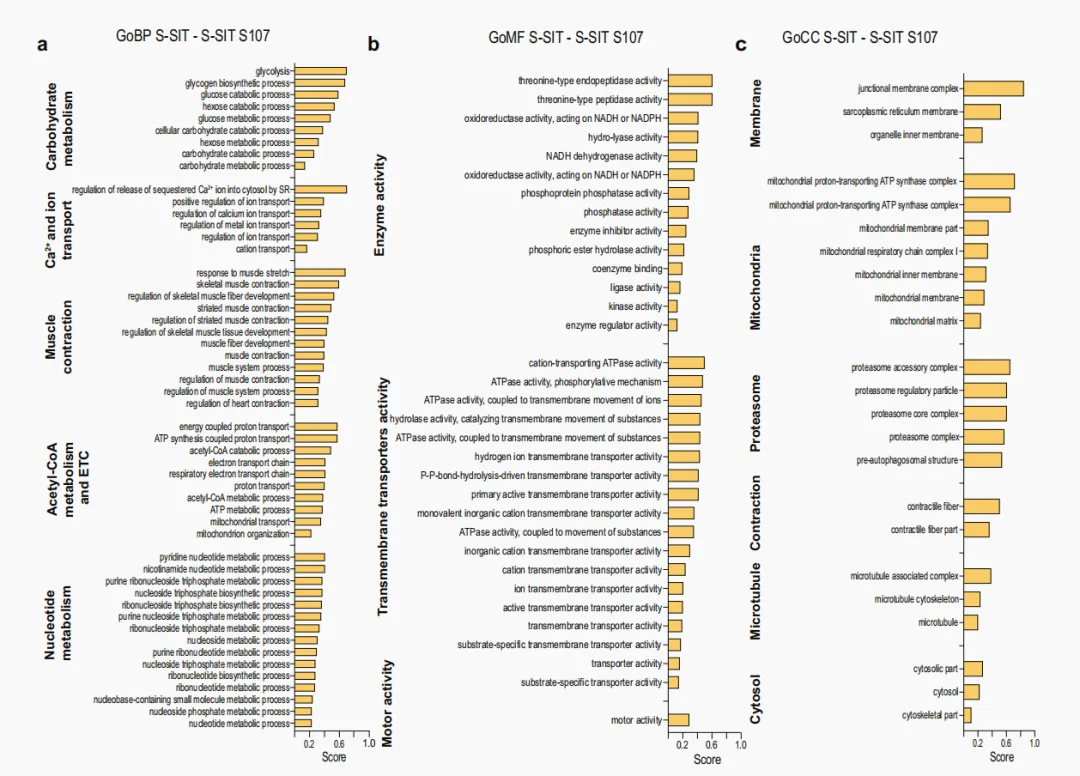
为进一步探究 RyR1 介导的急性 Ca²⁺ 泄漏所引起的有益肌肉代谢适应,对接受 S-SIT 处理的肌管在电刺激后分别给予或不给予 S107 处理 72 h,随后收集样本并用于蛋白质组学分析。将丰度发生显著变化的蛋白进行通路分析。经 S-SIT S107 处理的肌管中,与 DNA 和 RNA 加工相关的蛋白水平升高(补充图 3a–c);而以下生物学过程相关蛋白则显著下降:包括糖酵解在内的碳水化合物代谢、Ca²⁺ 及其他离子转运、肌肉收缩和肌纤维发育,以及包括 ETS 在内的线粒体功能(图 4a–c)。葡萄糖是 SIT 过程中使用的主要能量来源⁶⁰。葡萄糖可转化为丙酮酸,后者进一步转化为乳酸,或转化为乙酰辅酶 A(acetyl-coenzyme A,acetyl-CoA)后进入三羧酸(tricarboxylic acid,TCA)循环,从而进行充分的有氧代谢。鉴于 S107 处理是在电刺激后施加于 S-SIT 肌管的,S-SIT S107 肌管中特定通路相关蛋白整体下降提示,在 S-SIT 反应中,经 RyR1 发生的 Ca²⁺ 漏可触发骨骼肌的多种生理适应,其中碳水化合物代谢尤为突出。抑制 RyR1 介导的 Ca²⁺ 泄漏也可能降低 SERCA 介导的 Ca²⁺ 摄取以及胞质中的 ATP 利用,这可能成为影响我们在 S-SIT 和 S-SIT S107 肌管中观察到的线粒体变化的混杂因素。肌脂蛋白 (sarcolipin,SLN) 是一种小分子,已知可与 SERCA 结合并抑制其活性。SERCA 利用 ATP 水解释放的能量将 Ca²⁺ 跨 SR 膜转运⁶¹。SLN 与 SERCA 结合可促进 SERCA 泵的解偶联,使 Ca²⁺ 泄漏至胞质,而非进入 SR 腔内⁶²。刺激后 72 h,SIT 与 S-SIT S107 肌管之间 SERCA1(C2C12 肌管中表达的 SERCA 主要亚型⁶³)及其在骨骼肌中的主要调节因子 SLN⁶⁴的表达无显著差异(补充图 4b、c)。SERCA/SLN 免疫共沉淀实验显示,与对照组相比,S-SIT 和 S-SIT S107 肌管之间无显著差异(补充图 4a、d)。然而,已有研究表明,SLN 可降低 SERCA 对 Ca²⁺ 的摄取,但并不改变 ATP 水解⁶⁵,因此提示 SLN 可作为 SERCA 的解偶联因子发挥作用,即 SERCA 仍持续水解 ATP,但转运至 SR 腔内的 Ca²⁺ 减少⁶⁶。因此,我们制备了微粒体(富含 SR 的组分)(补充图 4e、f),以直接检测 SERCA1 的 ATP 酶活性。结果显示,在刺激后 72 h,与对照组和 S-SIT S107 肌管相比,S-SIT 肌管中的 ATP 酶活性升高(补充图 4g、h),提示 S-SIT 肌管 SR 中的 ATP 利用增加。值得注意的是,与对照组相比,S-SIT 和 S-SIT S107 肌管全细胞裂解液中的 ATP 水平均呈相似程度升高(补充图 4i)。尽管 S-SIT 肌管 SR 中的 ATP 利用增加,但其 ATP 水平仍较高,这提示与对照组和 S-SIT S107 肌管相比,S-SIT 肌管处于更高的代谢状态。
图 4|蛋白质组学分析显示 RyR1 Ca²⁺ 泄漏对 S-SIT 诱导肌肉适应的全局映射a–c 与 GoBP、GoMF 和 GoCC 相关的蛋白组蛋白质组学分析,显示在 S-SIT 后经 10 µM S107 处理 72 h 显著降低的蛋白组(处理在刺激后立即进行)。超过 400 个蛋白的蛋白组被排除。计算 S-SIT − S-SIT S107 差异的中位数,并根据差异幅度赋予评分。正评分表示被 S107 处理显著抑制的通路。每组 n = 5。采用 Benjamini–Hochberg 校正 t 检验。源数据作为 Source Data 文件提供。补充图 3|S107 处理增加 S-SIT 肌管中蛋白组的蛋白质组学数据a–c 与 GoBP(a)、GoMF(b)和 GoCC(c)相关的蛋白组蛋白质组学分析,这些蛋白组在 10 µM S107 处理 72 h 后显著增加(处理在刺激后立即进行)。超过 400 个蛋白的蛋白组被排除。计算 S-SIT − S-SIT S107 差异的中位数,并根据差异幅度赋予评分。负评分表示经 S107 处理显著增加的通路。每组 n = 5。Benjamini 校正 t 检验。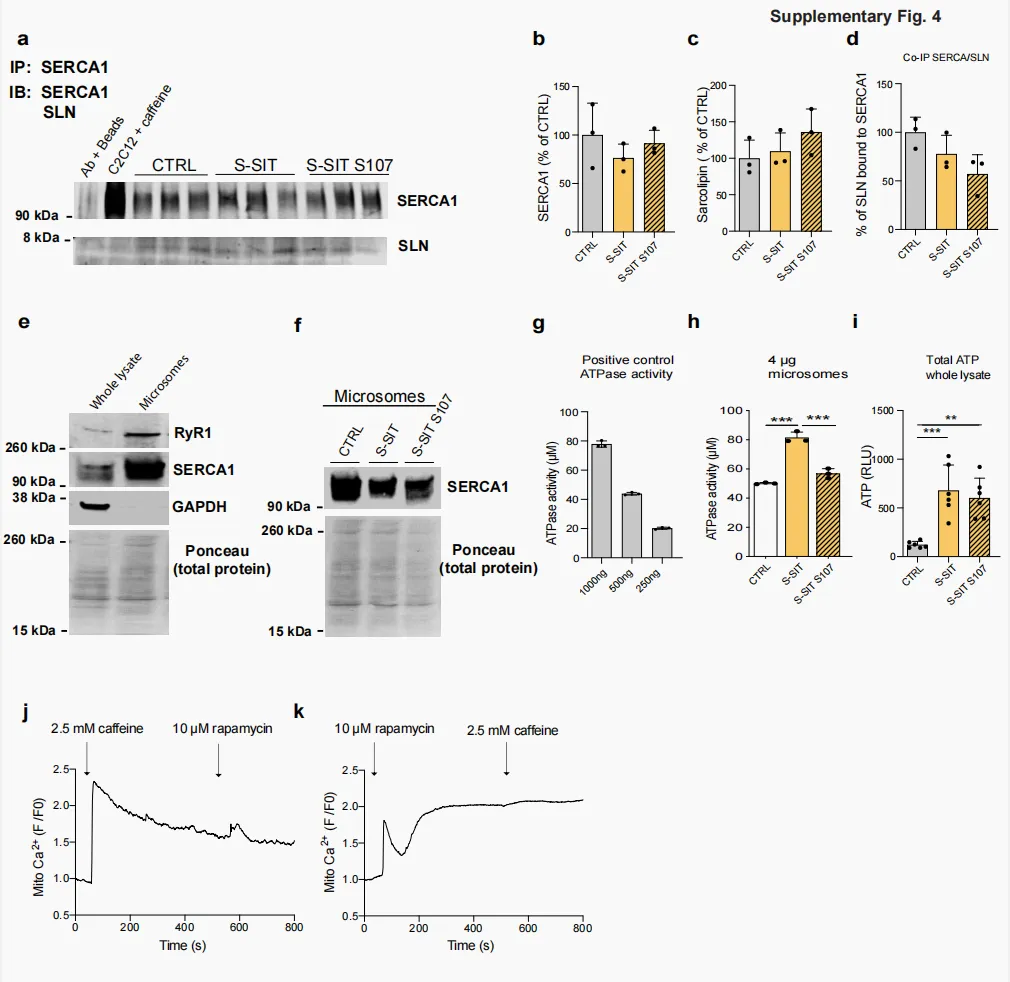 补充图 4|C2C12 肌管中 RyR1 开放诱导的线粒体 Ca²⁺ 摄取,以及 S-SIT 应答下肌管中 SERCA 活性的改变
补充图 4|C2C12 肌管中 RyR1 开放诱导的线粒体 Ca²⁺ 摄取,以及 S-SIT 应答下肌管中 SERCA 活性的改变a 刺激后 72 h,在未进行或进行 10 µM S107 处理的肌管中,IP SERCA1 的代表性 IB,以及 SERCA1和 SLN 的检测。b,d 对 CTRL、S-SIT 和 S-SIT S107 肌管中,输入样品中的 SERCA1(b)、SLN(c)以及结合到 SERCA1 上的 SLN 含量(d)进行定量分析;每组 n = 3 独立生物学实验。单因素ANOVA。e 微粒体制备的特征。与全裂解液相比,微粒体(SR 制备物)中 RyR1、SERCA1 和 GAPDH 的 IB。f CTRL、S-SIT 和 S-SIT S107 肌管中 SERCA-1 的代表性 IB(刺激和处理后 72 h)。g 阳性对照裂解液(全裂解液)中 ATP 酶活性的剂量反应定量,裂解液用量为 1000、500 和 250 ng。h 刺激和处理后 72 h,CTRL、S-SIT 和 S-SIT S107 肌管中 ATP 酶活性的定量;每组 n = 3 独立生物学实验。单因素 ANOVA,并进行 Tukey 多重比较检验。i 刺激和处理后 72 h,CTRL、S-SIT 和 S-SIT S107 肌管全细胞裂解液中 ATP 生成的定量。RLU;n = 3 独立生物学实验。单因素ANOVA,并进行 Tukey 多重比较检验。j C2C12 肌管中的标准化 Rhod-2 荧光成像,显示响应 2.5 mM 咖啡因刺激后再给予 10 µM 雷帕霉素时的线粒体 Ca²⁺ 摄取。箭头表示处理施加的时间点。k C2C12 肌管中的标准化 Rhod-2 荧光成像,显示响应 10 µM 雷帕霉素后再给予 2.5 mM 咖啡因时的线粒体 Ca²⁺ 摄取。箭头表示处理施加的时间点。S107 处理在刺激后立即进行,并持续 72 h。数据以mean ± SD 表示。**p ≤ 0.01,***p ≤ 0.001。源数据作为 Source Data 文件提供。
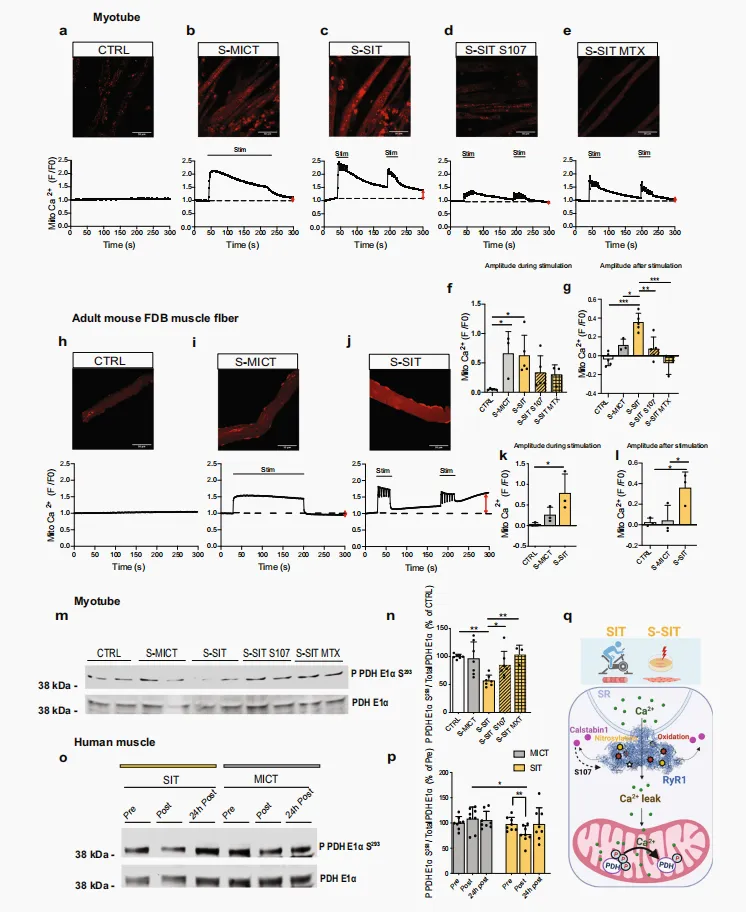
Ca²⁺ 是众所周知的第二信使,参与包括骨骼肌在内的多种组织的多种生理过程⁶⁷˒⁶⁸。经 SR 释放的 Ca²⁺ 可通过线粒体外膜上的电压依赖性阴离子通道进入线粒体⁶⁹˒⁷⁰,并进一步经 MCU 跨越线粒体内膜⁷¹。已有研究表明,这种线粒体 Ca²⁺ 流动与线粒体生物能量学的改善有关⁷²。因此,我们研究了 S-SIT 反应中经 RyR1 发生的 Ca²⁺ 泄漏是否会诱导线粒体 Ca²⁺ 摄取,从而触发有益的线粒体适应。我们采用 Rhod-2/AM 探针和共聚焦活细胞时间序列成像技术,检测单次 S-SIT 或 S-MICT 后的线粒体 Ca²⁺ 摄取情况¹⁴。负载 Rhod-2/AM 探针的对照肌管在光照暴露下未显示线粒体 Ca²⁺ 增加(图 5a),而S-MICT 和 S-SIT 肌管中的线粒体 Ca²⁺ 均显著增加(图 5b、c、f)。S-MICT 期间线粒体 Ca²⁺ 迅速恢复至基础水平,而 S-SIT 后则持续升高(图 5b、c、g),提示 S-SIT 后线粒体内 Ca²⁺ 发生积累。经 10 μM S107 预处理的 S-SIT 肌管未显示线粒体 Ca²⁺ 积累(图 5d),表明经 RyR1 发生的 Ca²⁺ 泄漏是 S-SIT 后观察到的线粒体 Ca²⁺ 摄取的主要来源。S-SIT 肌管经 20 μM MCU 抑制剂米托蒽醌 (mitoxantrone,MTX)⁷³ 预处理后,也获得了类似结果(图 5e、g)。这些结果提示,经 RyR1 泄漏的部分 Ca²⁺ 通过 MCU 进入线粒体,这支持了在单次 SIT 后即刻观察到的 SR Ca²⁺ 下降结果(图 2g–j)。
为确认 S-SIT 是否可在成熟骨骼肌中诱导线粒体 Ca²⁺ 积累,我们在分离的小鼠 FDB 肌纤维中检测了线粒体 Ca²⁺ 摄取。我们对 FDB 肌纤维施加 S-MICT 和 S-SIT,并参照 C2C12 肌管中的实验方法,在刺激结束后追踪线粒体 Ca²⁺ 的积累情况。对照组肌纤维在光照暴露下未显示线粒体 Ca²⁺ 增加(图 5h)。与 C2C12 肌管中的结果相似,S-MICT 和 S-SIT 均导致 FDB 肌纤维中的线粒体 Ca²⁺ 瞬变发生改变(图 5i、j、k)。同样,在 S-SIT 过程结束时,线粒体摄取的 Ca²⁺ 量高于 S-MICT 过程结束时的水平(图 5l)。
随后,我们在体外模型中检测了 RyR1 Ca²⁺ 泄漏是否足以形成局部高 Ca²⁺ 微区,从而通过 MCU 触发线粒体 Ca²⁺ 摄取。已知线粒体 Ca²⁺ 摄取发生于特定条件下,即 SR与线粒体之间需形成局部 Ca²⁺ 浓度升高的微区⁷⁴。尽管普遍认为 MCU 需要微摩尔浓度的 Ca²⁺ 才能激活,但也有证据表明线粒体在纳摩尔浓度下也能摄取 Ca²⁺ ⁷⁵˒⁷⁶。我们研究假设:在 RyR1 泄漏的响应下会形成这样的 Ca²⁺ 微区,从而触发线粒体 Ca²⁺ 摄取。我们首先在肌管中检测 RyR1 开放诱导的线粒体 Ca²⁺ 积累模式,通过使用 2.5 mM 咖啡因处理肌管,同时用 Rhod-2/AM 探针监测线粒体 Ca²⁺ 含量。咖啡因诱导了快速且大幅度的线粒体 Ca²⁺ 增加,随后逐渐下降,而加入 10 μM 雷帕霉素(可使 calstabin1 从 RyR1 解离¹⁴)则诱导了少量额外的线粒体 Ca²⁺ 摄取(补充图 4j)。当肌管先用 10 μM 雷帕霉素处理时,它们也表现出快速的线粒体 Ca²⁺ 增加,但幅度低于咖啡因处理(补充图 4k)。这种线粒体 Ca²⁺ 摄取呈逐渐增加并达到平台期,随后加入咖啡因并未引起进一步的线粒体 Ca²⁺ 增加(补充图 4k)。这些结果表明,雷帕霉素诱导的 RyR1 Ca²⁺ 泄漏足以引发渐进且持续的线粒体 Ca²⁺ 摄取。总体而言,在 C2C12 肌管细胞和成年小鼠 FDB 肌纤维中的观察表明,在 S-SIT 刺激下,RyR1 通道发生泄漏,导致线粒体摄取 Ca²⁺。虽然已知电刺激会引起线粒体 Ca²⁺ 摄取,但在电刺激结束后的 S-SIT 中观察到特异性的线粒体 Ca²⁺ 积累,这表明其机制是 RyR1 诱导的 Ca²⁺ 泄漏,而非电刺激诱发的 Ca²⁺ 释放。
图 5|(S-)SIT 诱导的 RyR1 泄漏增加肌细胞线粒体 Ca²⁺ 摄取,并降低 PDH 磷酸化水平a–e C2C12 肌管中的标准化 Rhod-2 荧光成像。对略作修改的单次 S-SIT 和 S-MICT 方案进行 300s 延时共聚焦记录:b S-MICT 和 c S-SIT。d,e 预先使用 10 µM S107 处理 1 h(d)或 20 µM米托蒽醌(e)的 S-SIT 肌管中的线粒体 Ca²⁺ 摄取。水平黑线表示刺激时间段;垂直红色箭头表示记录结束时线粒体 Ca²⁺ 的幅度。f–g(a–e)中刺激期间(f)以及 S-MICT 和 S-SIT 刺激结束后 80 s(g)标准化 Rhod-2 荧光的最大幅度;n = 5(CTRL、S-SIT 和 S-SIT S107)和 3(S-MICT、S-SIT MTX)个独立生物学实验。单因素 ANOVA,并进行 Tukey 多重比较检验。h–j 小鼠 FDB 完整单肌纤维中的标准化 Rhod-2 荧光成像。方案与(a–c)所述相同,但电压为 40 V。k–l (h–j)中 FDB 肌纤维在 S-MICT 和 S-SIT 刺激期间(k)以及刺激结束后 80 s(l)标准化 Rhod-2 荧光的最大幅度;每组 n = 3 只小鼠。单因素 ANOVA,并进行 Tukey 多重比较检验。m–n 肌管刺激后 1 h,Ser293 位点磷酸化 PDH E1α 相对于总 PDH E1α 的代表性 IB(m)及定量(n);如图中所示,刺激后给予 10 µM S107 或 20 µM 米托蒽醌处理 1 h;n = 7(CTRL、S-MICT、S-SIT 和 S-SIT S107)和3(S-SIT MTX)个独立生物学实验。单因素 ANOVA,并进行 Sidak 多重比较检验。o,p 人肌肉中Ser293 位点磷酸化 PDH E1α 相对于总 PDH E1α 的代表性 IB(o)及定量(p);每组 n = 8。双因素ANOVA,并进行 Sidak 多重比较检验。q S-SIT 应答中 RyR1 Ca²⁺ 泄漏激活线粒体 PDH E1α 去磷酸化的拟议示意图。数据以 mean ± SD 表示。*p ≤ 0.05,**p ≤ 0.01,***p ≤ 0.001。源数据作为Source Data 文件提供。 RyR1 Ca²⁺ 泄漏诱导的线粒体 Ca²⁺ 摄取使 PDH 去磷酸化,并构成 S-SIT 诱导 OXPHOS CI 增加的基础
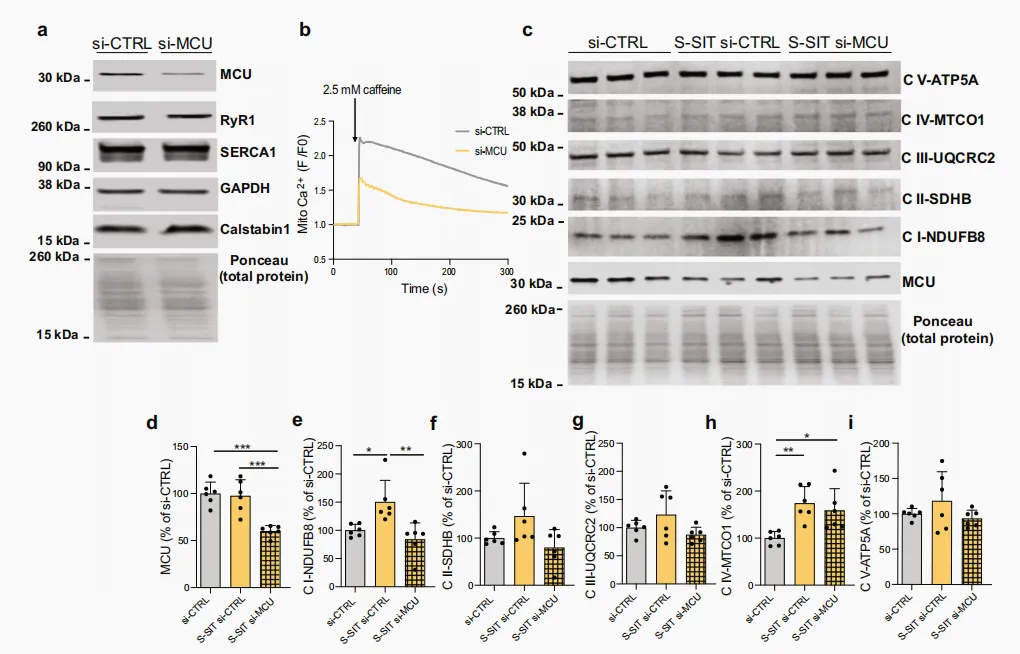
多种线粒体酶对 Ca²⁺ 敏感。其中,PDH 是碳水化合物来源的丙酮酸进入三羧酸循环进行完全氧化的关键门控酶⁷⁷。PDH 催化丙酮酸脱羧生成 acetyl-CoA,该反应伴随 NAD⁺ 还原为 NADH。PDH 活性受 PDH E1 亚基上四个残基(Ser293、Ser295、Ser300 和 Ser232)磷酸化的共价调控。PDH 激酶通过磷酸化使 PDH 失活,去磷酸化使 PDH 激活⁷⁸。在人类骨骼肌中,PDH 活性可随运动调节⁷⁹。以下原因促使我们研究 SIT 刺激下的 PDH 磷酸化:(i)骨骼肌 PDH 活性依赖运动强度⁸⁰,MCU−/− 小鼠在进行高强度运动时能力显著下降⁸¹;(ii)MCU 敲除会导致 PDH 磷酸化水平升高,其作用类似于 PDH 磷酸酶敲除,提示线粒体 Ca²⁺ 摄取与 PDH 磷酸化水平相关⁸²。因此,我们在 S-SIT 肌管中测定 PDH 磷酸化水平,以反映线粒体 Ca²⁺ 摄取的增加情况。在刺激后 1 小时,S-SIT 肌管中 PDH Ser293 磷酸化水平(P PDH E1α Ser293)显著下降,而 S-MICT 肌管未见明显变化(图 5m、n),反映了 SIT 诱导的线粒体 Ca²⁺ 摄取及 PDH 激活。在人类肌肉活检样本中也观察到 PDH 去磷酸化,仅在 SIT 后而非 MICT 后发生(图 5o、p),再次说明 S-SIT 模型可模拟单次 SIT 对人类骨骼肌的影响。为探讨 RyR1 Ca²⁺ 泄漏在该过程中可能的作用,我们在刺激后立即用 10 μM S107 处理 S-SIT 肌管 1 小时,然后测定 PDH 磷酸化水平。PDH 磷酸化下降被明显抑制(图 5m、n),这反映了 S-SIT 可诱导线粒体 Ca²⁺ 摄取和 PDH 激活。S-SIT 肌管在刺激后立即用 20 μM MTX 处理 1 小时,也可恢复 PDH 磷酸化水平,进一步确认线粒体 Ca²⁺ 摄取在该过程中的作用(图 5m、n)。综上所述,SIT 可诱导 RyR1 翻译后修饰及 calstabin1 从 RyR1 解离,导致 RyR1 泄漏、线粒体 Ca²⁺ 摄取增加以及 PDH 去磷酸化(图 5q)。我们进一步研究了 RyR1 Ca²⁺ 泄漏诱导的线粒体 Ca²⁺ 摄取是否在 S-SIT 后较长时间(刺激后 72 h)的线粒体适应中发挥作用。为此,在刺激后立即用靶向 MCU 的小干扰 RNA(siRNA)转染 S-SIT 肌管细胞,并在 72 h 后测定 OXPHOS 蛋白。我们首先确认了 si-MCU 能够有效降低分化后肌管细胞中 MCU 的表达和线粒体 Ca²⁺ 摄取(图 6a、b),同时不影响参与 SR Ca²⁺ 瞬变的主要蛋白(RyR1、SERCA 和 calstabin1)的表达(图 6a)。与转染 siRNA 的对照组(si-CTRL 和 S-SIT si-CTRL)相比,S-SIT 刺激后立即转染 si-MCU 的肌管细胞,在处理 72 h 后表现出 MCU 蛋白水平降低(图 6d)。S-SIT si-CTRL 肌管的 OXPHOS CI 和 CIV 蛋白显著上调,而 S-SIT si-MCU 则特异性抑制了 OXPHOS CI 的增加(图 6c、e–i)。由于是在 S-SIT 之后下调 MCU 表达,因此 S-SIT 介导的线粒体 Ca²⁺ 摄取是导致线粒体 CI 表达增加的原因。这一发现与近期研究一致,该研究显示 Barth 综合征患者心脏组织中 MCU 水平降低与 OXPHOS CI 蛋白特异性下降相关⁸³。综上,这些观察结果揭示了 MCU 及线粒体 Ca²⁺ 在调控 OXPHOS CI 中的重要作用,为进一步研究提供了新的方向。图 6|线粒体 Ca²⁺ 摄取促进肌管对 S-SIT 的线粒体适应a si-MCU(针对 MCU 的 siRNA)与 si-CTRL(阴性对照 siRNA)肌管中 MCU、RyR1、SERCA1、GAPDH 和 calstabin1 的免疫印迹(IB)。b si-MCU 与 si-CTRL 肌管在转染后 72 h 的标准化 Rhod-2 荧光成像原始记录。c S-SIT si-MCU 肌管在刺激后 72 h(刺激后立即转染针对 MCU 的 siRNA)线粒体 OXPHOS 和 MCU 蛋白表达的代表性 IB,与 S-SIT si-CTRL 肌管(刺激后立即转染阴性对照 siRNA)及 si-CTRL(未刺激、转染阴性对照 siRNA 的肌管)相比。所有 OXPHOS 蛋白裁剪部分来自同一胶,完整胶见补充图6。d–i (c)中免疫印迹相对于总蛋白的定量,并表示为 si-CTRL百分比;每组 n = 6 独立生物学实验。单因素 ANOVA 并进行 Tukey 多重比较检验(d,e,h)。数据以 mean ± SD 表示。*p ≤ 0.05,**p ≤ 0.01,***p ≤ 0.001。源数据作为 Source Data 文件提供。本研究发现,单次 SIT 而非 MICT 可诱导人类骨骼肌 RyR1 发生 PTMs,并继而导致 calstabin1 解离;类似地,在接受 S-SIT 而非 S-MICT 处理的 C2C12 肌管中亦观察到上述变化。上述现象与人类 SIT 后骨骼肌及 S-SIT 肌管中更为显著的线粒体适应性改变相一致。基于体外模型,本研究进一步从因果层面证实,RyR1 Ca²⁺ 泄漏与线粒体重塑及线粒体功能改善密切相关;这一判断的依据在于,RyR 稳定剂 S107 可显著减弱上述线粒体重塑及功能改善效应。因此,在健康个体中,高强度运动诱导的骨骼肌急性 RyR1 Ca²⁺ 泄漏可被视为一种具有适应性意义的生理反应,其原因在于该过程能够促进线粒体相关蛋白水平升高。该发现进一步表明,健康状态下的急性 RyR Ca²⁺ 泄漏应与病理状态下持续性 RyR Ca²⁺ 泄漏应加以区分。既往研究中,我们报道了人类单次 SIT 后 RyR1 出现片段化,可能导致 RyR1 通道发生泄漏²⁶。后续研究表明,RyR1 片段化依赖受试者的易感性及训练状态,由此提出了“响应者”与“非响应者”的概念²⁸。本研究重点关注 calstabin1 从 RyR1 解离的现象,因为这是 RyR1 通道泄漏的关键标志⁵³。我们的研究假设为:SIT 诱导的急性 RyR1 Ca²⁺ 泄漏可引发骨骼肌线粒体重塑,并改善线粒体呼吸功能。我们的结果从机制层面显示,S-SIT 诱导 RyR1 泄漏的 Ca²⁺ 可被线粒体摄取,并触发 PDH 去磷酸化,其证据包括:(i)与 S-MICT 肌管相比,S-SIT 肌管线粒体 Ca²⁺ 积累更为显著,而 S107 处理可消除该表型;(ii)与 S-MICT 和 MICT 相比,S-SIT 肌管和人类 SIT 骨骼肌中 PDH 磷酸化水平显著下降;(iii)S107 处理以及 MTX 对 MCU 的抑制均可恢复 S-SIT 肌管中 PDH 磷酸化水平。在骨骼肌中,SR 与线粒体在空间上紧密耦联⁶⁹,SR 释放的 Ca²⁺ 可被线粒体摄取⁷²˒⁸⁴˒⁸⁵。肌细胞去极化可增强线粒体 Ca²⁺ 摄取,该过程与线粒体生物能量代谢密切相关,因为肌细胞去极化过程中抑制 RyR1 会降低 ATP 依赖性的 O₂ 消耗⁷²。在我们的 S-MICT 和 S-SIT 模型中,电刺激期间线粒体 Ca²⁺ 水平均升高,并在刺激结束后下降。S-MICT 肌管中线粒体 Ca²⁺ 几乎完全恢复至基线,而 S-SIT 肌管则表现出小幅但持续存在的线粒体 Ca²⁺ 平台期,该现象与肌细胞去极化无直接关联,并可能激活线粒体 Ca²⁺ 依赖性酶 PDH。S-SIT 后早期 PDH 去磷酸化虽不能直接与后期线粒体适应性改变(72 h 后 OXPHOS 蛋白及呼吸增加)建立联系,尤其是在底物饱和条件下的呼吸测定中,但以下证据支持 RyR1 Ca²⁺ 泄漏在 NADH 依赖性线粒体适应性中的作用:(i)蛋白质组学数据显示 S107 处理可改变与 PDH 活性相关的蛋白组(补充数据 1 和 2);(ii)S-SIT S107 肌管 NADH 水平下降。肌管中 MCU 下调可削弱 S-SIT 诱导的 OXPHOS CI 蛋白(NDUFB8)升高,也提示 RyR1 Ca²⁺ 泄漏在 S-SIT 诱导的线粒体适应性改变中具有特异性作用。综合上述结果及 S-SIT S107 的数据,支持了 RyR1 Ca²⁺ 泄漏及线粒体 Ca²⁺ 摄取在 S-SIT 诱导的线粒体适应性改变中发挥有益调控作用。Jain 等⁸⁶既往研究表明,高脂饮食可增加线粒体 ROS 生成,进而触发肌肉细胞中 RyR1 的 S-亚硝基化及 CaMK 磷酸化,并伴随线粒体生物发生增强。本研究发现,SIT 与 MICT 诱导的 RyR1 PTMs 模式存在差异,但二者引起的 CaMK 磷酸化水平相近。在骨骼肌收缩过程中,SR 快速释放大量 Ca²⁺,可迅速且显著诱导 CaMK 磷酸化并增强其活性⁸⁷,这解释了 SIT 与 MICT 中 CaMKII 磷酸化水平均呈相似升高的现象。此外,Wright 等³⁸曾报道,细胞质 Ca²⁺ 浓度升高可通过 CaMK 依赖性途径激活 p38,并促进 PGC-1α 表达;这一发现亦支持本研究结果,即 SIT 与 MICT 均可诱导 CaMKII 磷酸化水平及 PGC-1α 表达相似升高。尽管 SIT 与 MICT 在上述指标上表现出相似变化,本研究仍提示,单次 SIT 诱导的 RyR1 Ca²⁺ 泄漏所介导的线粒体 Ca²⁺ 摄取,与 NADH 依赖性线粒体适应性改变的激活之间存在特异性的因果关联。已有观点认为,HIIT 诱导的线粒体适应可能需要经过多日或多次运动才能产生显著的有益效应⁸⁸。近期研究中,Skelly 等报道,单次 SIT 和 MICT 均可在人类各种肌纤维中快速诱导线粒体生物发生标志物的变化,而长期训练则表现出肌纤维类型依赖性的差异性适应⁸⁹。Trewin 等³¹的独立研究显示,尽管 HIIT 的运动总量相对较低,但单次 HIIT 与 MICT 均可引起线粒体蛋白水平及线粒体呼吸功能改变。此外,与 MICT 相比,SIT 可迅速诱导 PGC-1α 蛋白发生核定位⁹⁰。在本研究中,SIT 与 MICT 均可使人类骨骼肌及细胞中 PGC-1α 蛋白水平升高,但仅 SIT 显著增加了 OXPHOS CI 和 CII 蛋白水平。考虑到 PGC-1α 的核定位对其功能正常发挥至关重要,这提示 SIT 与 MICT 在其功能调控机制上可能存在差异。此外,目前尚不清楚我们观察到的急性线粒体诱导效应能否转化为长期适应性改变。已有多项研究比较了数周 SIT 与 MICT 的代谢适应性,结果显示二者均可类似地提高有氧能力(VO₂max)和线粒体含量²³˒⁹¹˒⁹²。然而,线粒体生物发生所依赖的细胞应激及代谢信号被证明与运动强度密切相关;少量研究显示,对同一受试者进行等功比较时,SIT 可比 MICT 更显著地增加线粒体含量⁹¹。规律性运动可在多种细胞、组织及器官(如骨骼肌、心肌、大脑、肝脏及脂肪组织)中引起广泛的生理适应,从而带来多重健康益处¹⁷。对于健康的中年男性而言,身体健康水平是预测死亡风险的重要指标,身体健康的轻微改善与全因死亡风险的显著降低密切相关¹⁶。尽管当前指南表明,任何单次身体活动均可带来健康益处¹⁷,鉴于运动反应可能随个体体能状态而异,因此有必要明确,在运动类型、运动量和运动强度不同的时间高效型运动组合中,何种组合最有利于实现特定结局。为获得最大化健康获益,尤其是对于健康且体能良好的人群而言,即便这一目标需要通过规律性较高强度身体活动实现,其运动负荷也可能不宜过高⁹³。近期一项人体试验比较了 12 周 HIIT、抗阻训练及二者联合训练对心脏代谢健康指标和骨骼肌适应性改变的影响。结果显示,仅 HIIT 可改善心脏代谢健康指标,而 HIIT 和联合训练均可提高有氧能力及骨骼肌线粒体呼吸功能²²。蛋白质组学分析进一步显示,HIIT 训练后,ETS、OXPHOS、TCA 及糖酵解相关蛋白组水平均升高²²。采用最大强度间歇训练模式后,本研究发现,在单次 S-SIT 后使用 S107 抑制 RyR1 Ca²⁺ 泄漏,可降低 S-SIT 肌管中与糖酵解、acetyl coA 代谢、ETC、肌肉收缩及离子转运(包括 Ca²⁺ 转运)相关的蛋白组水平。这一结果表明,RyR1 Ca²⁺ 泄漏在触发运动诱导骨骼肌线粒体功能改善的机制中具有重要意义。由此提示,急性 RyR1 Ca²⁺ 泄漏不仅可诱导线粒体重塑,还可能在骨骼肌对运动训练产生的多种适应性改变中发挥关键调控作用。本研究仍存在一些局限性。为从机制层面阐明 RyR1 Ca²⁺ 泄漏与 SIT 诱导的骨骼肌有益适应之间的因果关联,本研究构建了体外 SIT 和 MICT 模型,并将其所得数据与人体观察结果相结合。鉴于本研究重点关注 RyR1 介导的 Ca²⁺ 泄漏相关线粒体适应,体外模型的设计旨在进一步明确人体骨骼肌中观察到的主要变化。尽管上述模型的结合有助于解析 RyR1 Ca²⁺ 泄漏所触发的线粒体适应机制,但本研究所建立的体外模型尚不能充分代表人体实际 SIT 和 MICT 模式。此外,因为 Ca²⁺ 可能并非该过程中的唯一作用机制,所以本研究结果并不能排除其他机制参与上述适应性改变的可能性。综上所述,SIT 是一种省时高效的运动干预方式,可有效抵消身体活动不足带来的不良影响,而身体活动不足是全球范围内主要的健康风险因素之一。本研究深入探讨了 SIT 对骨骼肌产生有益作用的分子机制,得出两点主要结论:(i)高强度运动可诱导 RyR1 发生 PTMs,使其进入急性泄漏状态;(ii)这一急性泄漏与骨骼肌线粒体重塑及功能改善之间存在因果关联。在概念层面上,我们认为,骨骼肌中的急性 RyR1 Ca²⁺ 泄漏可被视为有益的生理反应,并且与病理状态下观察到的持续性 RyR Ca²⁺ 泄漏存在显著差异。这些结论为进一步研究急性 RyR Ca²⁺ 泄漏在其他生理条件下的作用提供了理论基础。然而,急性 RyR Ca²⁺ 泄漏在病理状态下是否同样具有有益作用,仍有待进一步验证。