



本期信熹研究聚焦表观遗传编辑技术,系统梳理其概念、发展脉络与技术特点,进而探讨在功能基因组学和细胞工程的应用前景,同时分析当前面临的技术与临床应用挑战,介绍领域内的代表性企业。
一、表观遗传学和表观遗传编辑
1、 表现遗传调控机制
人体内所有细胞几乎拥有相同的基因编码,但细胞表型和功能却呈现出丰富的多样性,这是由于细胞内存在高度协调的、细胞类型特异性的基因调控。这种基因调控通过化学修饰来控制同一套基因组的表达,即表观遗传调控。
表观遗传学(Epigenetics)是研究DNA和组蛋白的生物化学修饰等方式如何调控基因表达模式的一门学科,这些修饰是可逆的、可遗传的,并在细胞分裂过程中持续存在,但不涉及DNA序列的改变。表观遗传修饰响应环境刺激(如外部信号或细胞环境变化),在发育、疾病发生、细胞记忆和表型可塑性中起关键作用,甚至介导跨代遗传的环境适应记忆。它通过多层次、动态的分子机制调控基因组功能,其核心机制包括DNA甲基化、组蛋白翻译后修饰(PTM)、染色质重塑、非编码RNA调控的相互作用。
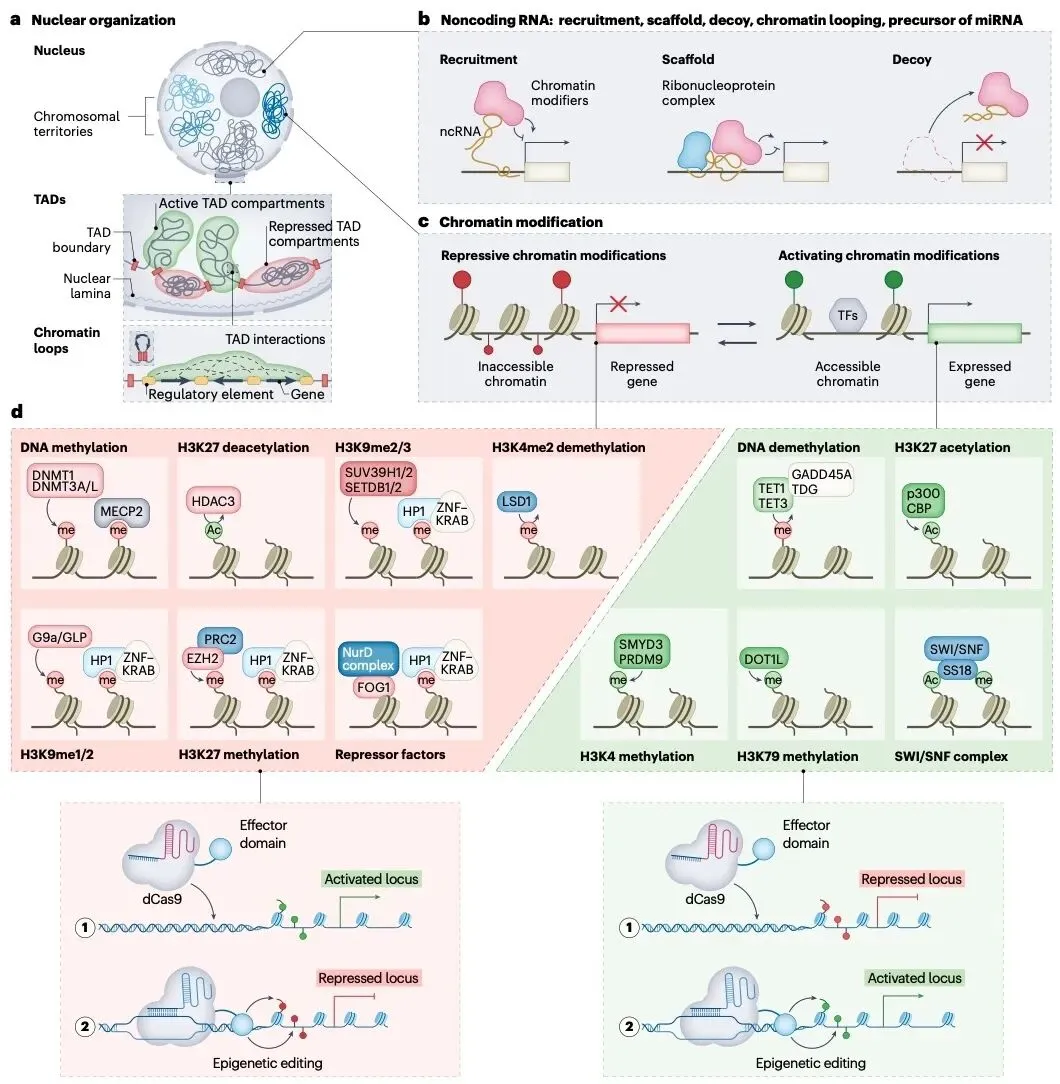
图1 | 表观遗传调控基因表达的示意图
(来源:Nat Biotechnol. 2024;42(8):1199-1217.)
DNA甲基化是由DNA甲基转移酶(DNMT)催化胞嘧啶C5位形成5-甲基胞嘧啶(5mC),主要富集于基因启动子CpG岛,通过空间阻碍转录因子结合或招募甲基结合蛋白(如MeCP2)形成抑制性复合物实现基因沉默。其可逆性由TET双加氧酶介导的氧化去甲基化途径实现,这一动态平衡响应发育信号与环境刺激。
组蛋白H3/H4尾部的赖氨酸、精氨酸残基受特异性酶系统调控,由所谓的“书写者”和“擦除者”引入和去除修饰基因,从而完成组蛋白翻译后修饰,并被染色质重塑剂识别。组蛋白甲基化是由组蛋白甲基转移酶(HMT)添加甲基基团,通过招募“阅读器”蛋白(如HP1结合H3K9me3)形成抑制性染色质;去甲基化酶(KDM)可移除甲基化修饰实现激活。组蛋白乙酰化是由组蛋白乙酰转移酶(HAT)添加乙酰基团中和组蛋白正电荷,松弛核小体结构促进转录;组蛋白去乙酰化酶(HDAC)可逆转该过程增强染色质压缩。此外,组蛋白还可以经历其他修饰,包括赖氨酸泛素化、赖氨酸类泛素化修饰、ADP-核糖基化、丝氨酸/苏氨酸/酪氨酸磷酸化等。
染色质重塑与核架构机制受多重因素调控。ATP依赖的染色质重塑复合物(如SWI/SNF)通过滑动、驱逐或替换核小体改变DNA可及性。染色质拓扑结构由染色质结构蛋白CTCF/黏连蛋白介导的拓扑关联结构域(TAD)组织,增强子-启动子环化受组蛋白修饰与结构蛋白协同调控。异染色质蛋白HP1通过相分离驱动异染色质区室化,维持基因沉默状态的物理隔离。
非编码RNA调控涉及多种类型的功能作用。长链非编码RNA(lncRNA)直接招募PRC2等修饰酶实现染色体剂量补偿。小干扰RNA(siRNA)通过RNA诱导沉默复合体(RISC)介导异染色质形成。增强子RNA(eRNA)与转录因子协同稳定增强子-启动子互作环路。
表观遗传调控构成了一个复杂的细胞过程网络,共同塑造并维持细胞的“表观遗传状态”,从而在基因组序列相同的情况下指导细胞功能的多样性。这一状态的形成和功能调控依赖于多种相互作用的因素,包括染色质可及性、DNA和组蛋白的修饰、与基因组相互作用的非编码RNA和蛋白质因子的活性,以及基因组三维结构和相分离过程等。这些调控因素并非独立运作,而是存在复杂的“串扰”(crosstalk),形成级联调控网络,在时空维度精确控制基因表达,例如组蛋白修饰与DNA甲基化之间会相互影响,共同决定最终的基因表达模式、细胞分化和功能记忆。执行这些表观遗传调控功能的内源性分子(主要是酶和蛋白复合物)被称为表观遗传效应因子(Epigenetic effectors),它们通过直接修饰染色质或介导结构变化来建立和维持特定的表观遗传状态。
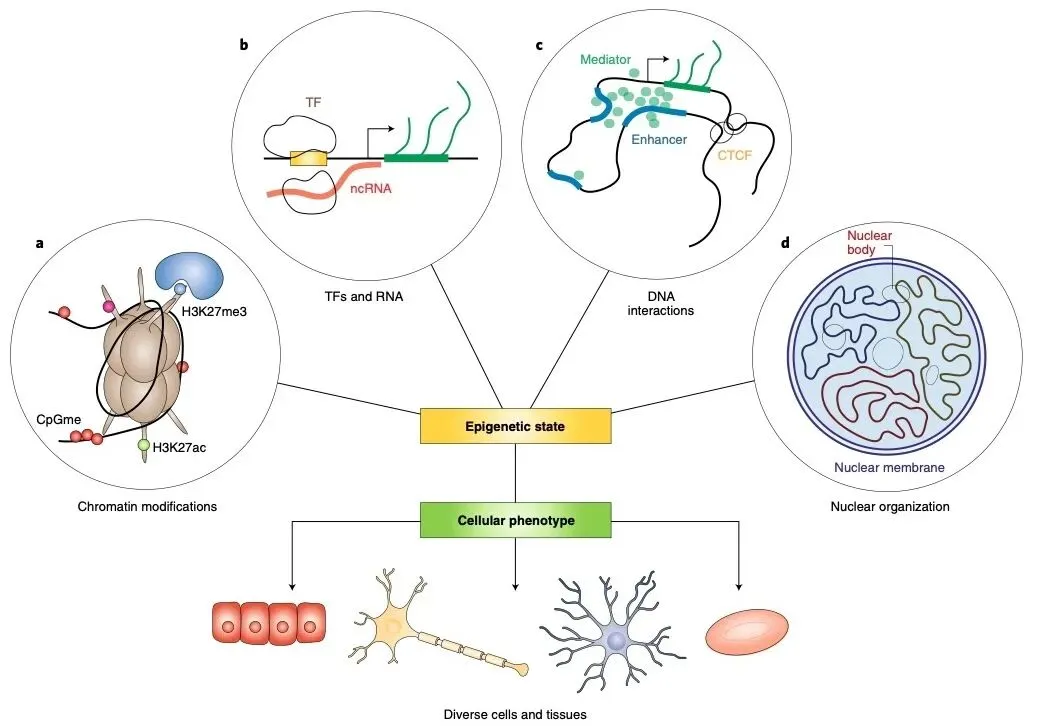
图2 | 细胞内的表观遗传过程
(来源:Nat Cell Biol. 2021;23(1):11-22.)
生物体对环境及药物刺激的易感性,以及几乎所有其他复杂表型如疾病和老化失调,主要是由基因调控的改变驱动的,而非基因本身的DNA序列变化。表观遗传修饰在不改变DNA序列的前提下,动态塑造细胞类型特异性的基因表达程序,从而确保了每个细胞在其生命周期内都有恰当的基因表达行为。其在生命过程中不仅指导发育编程与决定细胞命运,更通过响应环境刺激调控个体对疾病的易感性。表观遗传过程的扰动或动态失衡可能对健康和行为产生深远影响,导致成瘾、神经退行性疾病、自身免疫疾病和肿瘤等病理状态的发生。
2、针对表观遗传调控的药物开发
表观遗传失调在肿瘤疾病中屡见报道,这促进了表观遗传疗法在肿瘤领域的研究发展,并在血液系统恶性肿瘤治疗中取得了临床突破。迄今已有十余款涉及表观遗传的药物获批用于治疗肿瘤,其作用机制为DNA或组蛋白修饰酶抑制剂。FDA已批准两种针对DNMT的表观遗传药物——阿扎胞苷和地西他滨,用于治疗骨髓增生异常综合征。此外,四种HDAC抑制剂,即伏立诺他、罗米地辛、贝利司他和帕比司他,也已获FDA批准,分别用于治疗皮肤T细胞淋巴瘤(CTCL)、难治性CTCL、外周T细胞淋巴瘤(PTCL)和多发性骨髓瘤。
这些药物为非位点特异性全基因组表观遗传治疗药物,通过系统性干预表观遗传修饰酶的活性,实现对疾病相关基因表达网络的全局调控。然而,由于这些药物通常缺乏基因特异性,会导致全表观基因组修饰,有可能会增强抑癌基因沉默或上调促转移酶表达,从而促进肿瘤发生、进展和侵袭性。目前已获批的药物主要用于治疗血液瘤,而在实体瘤中的疗效受限,其原因一方面可能是由于血液淋巴系统恶性肿瘤的增殖率更高,而实体瘤细胞分裂相对缓慢,两者对药物的敏感性不同,另一方面药物缺乏特异性可能导致全基因组表观遗传修饰紊乱,增加药物的副作用。因此,基因组学将位点特异性表观遗传编辑作为下一种精准医学形式,以实现对特定基因组位点的精确编辑。
3、表观遗传编辑技术
表观遗传编辑技术是一种通过靶向修饰表观遗传标记(如DNA甲基化、组蛋白修饰)而非改变DNA序列本身来精确调控基因表达的生物技术。其核心工作原理依赖于工程化融合蛋白的定向招募:利用可编程DNA结合域(如锌指蛋白(ZFP)、转录激活样效应子(TALE)或失活Cas9蛋白(dCas9))特异性定位至目标基因组位点(如基因启动子或增强子),进而通过连接的效应结构域(Effector domain)在局部执行表观遗传重编程。这种“靶向+修饰”机制可实现基因的持久沉默或激活,例如通过诱导DNA甲基化或组蛋白甲基化实现转录抑制,或利用组蛋白乙酰化及DNA去甲基化激活基因表达。
表观遗传编辑工具的核心结构由两个功能模块协同构成,即靶向模块和效应模块。DNA靶向模块作为分子导航系统,其中dCas9通过sgRNA引导识别特定DNA序列,而ZFP和TALE则通过蛋白质-DNA相互作用直接结合目标位点。效应模块作为功能执行单元,包含表观遗传修饰酶(如甲基转移酶DNMT3A、去甲基化酶TET1、乙酰转移酶p300)或转录调控因子(如抑制域KRAB、激活域VP64),通过催化局部表观标记的写入或擦除改变染色质状态。该技术的独特优势在于其“hit-and-run”机制——短暂表达编辑器即可建立自我维持的表观遗传记忆,实现单次干预下的持久疗效。
二、表观遗传编辑技术的发展
表观遗传编辑工具的发展经历了从蛋白质工程到RNA编程的范式转变,其核心在于通过融合可编程DNA结合模块与表观效应结构域,实现对特异性位点的表观基因组重塑。早期ZFN技术利用人工设计的Cys2His2锌指结构域串联体识别DNA序列(每个锌指单元靶向3个碱基),通过融合表观效应结构域实现靶向修饰。然而,ZFN的构建复杂,需经验性筛选有效组合,且存在脱靶毒性风险。随后发展的TALE通过模块化重复单元(每个单元识别单个碱基,RVD决定特异性)提升了靶向灵活性,但其大分子量(>100 kDa)限制了递送效率。
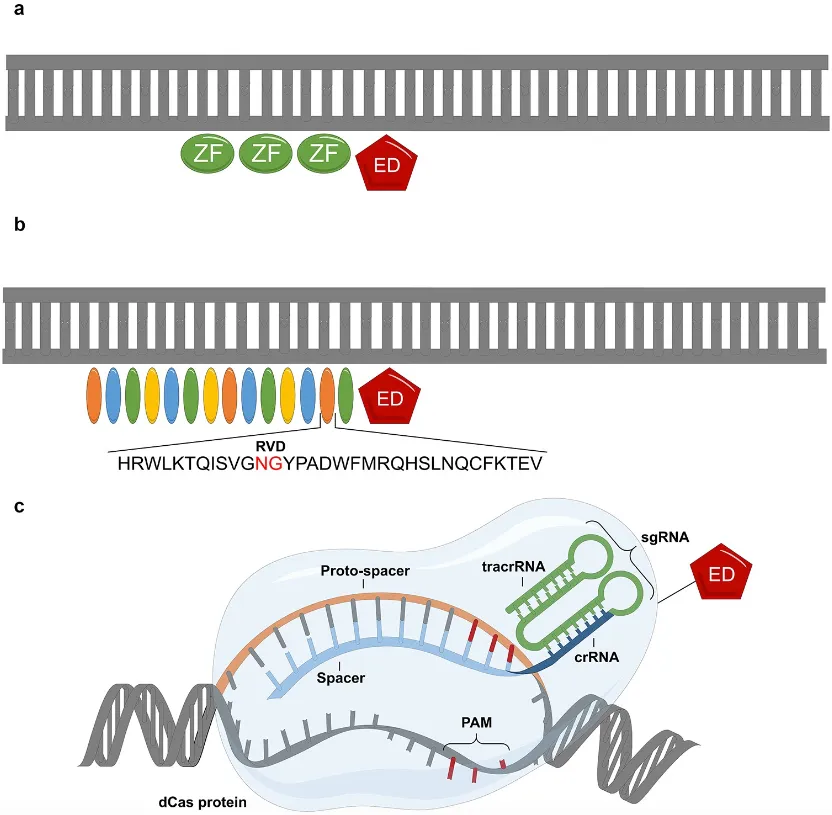
图3 | 表观遗传工具示意图
(来源:Epigenetics. 2022;17(5):564-588.)
CRISPR-dCas9系统的出现标志着技术拐点,dCas9与表观效应结构域的融合产生了一系列丰富的转录抑制和激活工具集(分别称为CRISPRi和CRISPRa)。dCas9核酸酶经突变后失去了切割DNA的能力,但仍保留通过sgRNA引导至靶DNA结合的活性,成为模块化表观编辑平台的基础。其核心优势在于sgRNA设计的简易性(仅需设计20个核苷酸长度的序列),但也存在PAM序列依赖性及染色质可及性的限制。为了优化调控效能,dCas9体系衍生出多代增强工具。
表1 | 重要表观遗传激活与抑制效应域
(来源:Nat Biotechnol. 2024;42(8):1199-1217.)
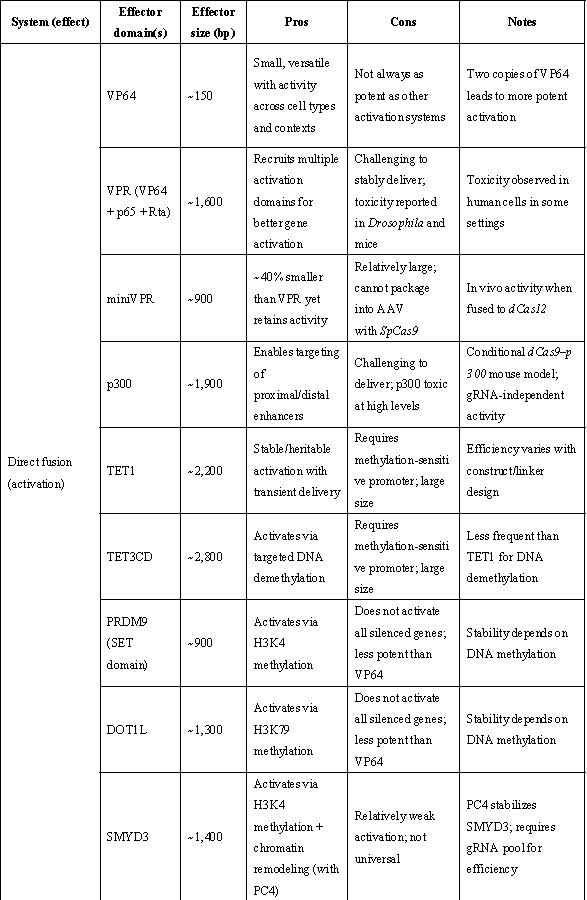
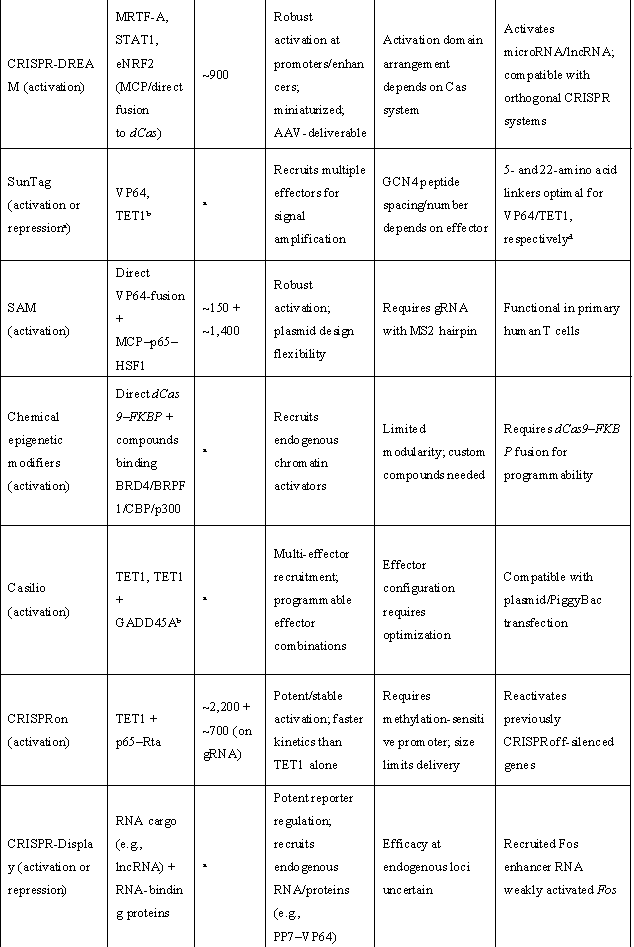
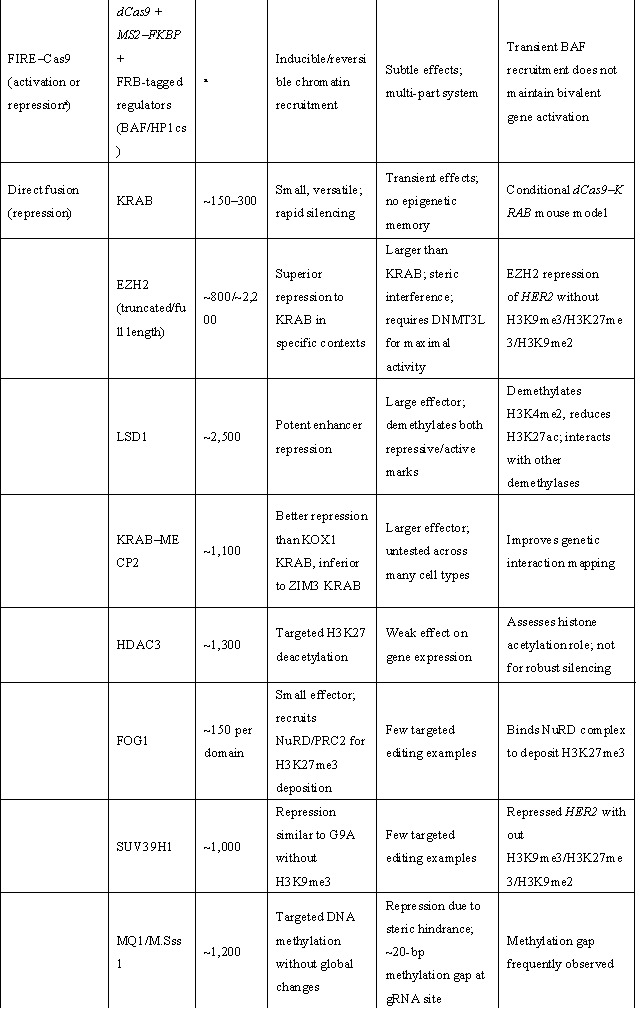
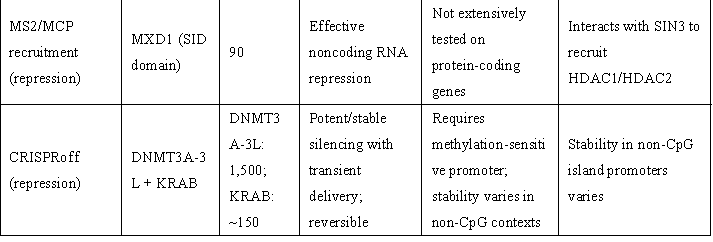
ᵃ 取决于招募的效应子。ᵇ 模块化(可使用其他效应子)。c 针对测试的连接子(linker)长度。
CRISPRa v1是将dCas9直接融合单一VP64激活域。VP64招募组蛋白乙酰转移酶及基础转录因子,通过局部组蛋白乙酰化松弛染色质结构,促进RNA聚合酶Ⅱ组装,实现基础转录激活。但单一VP64激活强度有限,对高甲基化或异染色质化区域无效。
CRISPRa v2 VPR系统是将VP64-p65-Rta三激活域串联融合至dCas9,显著提升激活强度。除了VP64的基础转录激活功能外,p65招募组蛋白乙酰化酶和染色质重塑复合物,增强启动子开放性,Rta通过结合RNA解旋酶复合物加速转录延伸。因此,融合p65与Rta可分别激活转录过程中自起始至延伸的不同阶段,发挥协同效应,使基因激活效率提升30-100倍。但VPR系统的分子量偏大(>5 kDa),增加了体内递送的难度,并且Rta结构域可能触发免疫应答。
CRISPRa v2 SunTag系统是将dCas9融合多拷贝GCN4肽阵列,同时招募多个scFv抗体-效应域融合蛋白,实现信号放大。其中,dCas9融合GCN4肽重复阵列作为支架蛋白,VP64或TET1等激活域与单链抗体scFv(抗GCN4)融合组成效应模块。单个dCas9-GCN4可通过亲合力作用招募10-24个scFv-VP64分子,形成局部高浓度效应域簇,强力激活转录或催化DNA去甲基化。但SunTag系统引入的scFv结构在哺乳动物细胞中表达稳定性较差,需对密码子优化。
CRISPRa v2 SAM系统是在sgRNA中插入MS2茎环结构,将MS2-p65-HSF1复合物招募到dCas9-VP64融合蛋白,增强激活特异性。SAM系统在dCas9-VP64融合蛋白的基础激活功能之外,对sgRNA改造插入了可与MS2衣壳蛋白特异性结合的MS2 RNA适配体茎环,并将MS2衣壳蛋白与p65和HSF1(热休克因子1)融合。其中,HSF1可招募组蛋白乙酰化酶及染色质解旋酶,协同VP64突破抑制性染色质屏障。SAM系统采用三重效应域(VP64+p65+HSF1)空间分离的架构,避免融合蛋白折叠冲突,激活效率较VPR可进一步提升。
CRISPRoff和CRISPRon系统代表了表观遗传编辑技术的重大突破,通过可编程的DNA甲基化与去甲基化实现基因表达的持久调控。CRISPRoff系统由dCas9融合KRAB抑制域、DNMT3A甲基转移酶及DNMT3L调节亚基三重效应域构成。其中,KRAB的作用是募集HP1/SETDB1复合物诱导组蛋白H3K9me3甲基化修饰建立抑制性的异染色质标记,DNMT3A的功能是催化CpG岛DNA甲基化,DNMT3L自身不具备催化活性,但可增强DNMT3A催化效能与靶向低甲基化染色质区域的特异性。早期研究发现染色质调控元件的抑制效果具有显著的异质性,其动力学特征、作用强度及稳定性高度依赖于目标基因的特定属性、细胞类型及局部染色质环境。KRAB通能迅速实现基因沉默,但其效应通常短暂且可逆,难以在多次细胞分裂中稳定维持。相比之下,DNMT3A/DNMT3B介导的从头DNA甲基化虽启动较慢且初始沉默强度较弱,却能通过建立稳定的CpG岛甲基化标记,形成持久的表观遗传记忆,使基因沉默状态在细胞增殖过程中可遗传。CRISPRoff通过建立DNA甲基化和沉积抑制性组蛋白修饰的协同作用,不仅实现了对多种内源基因的强效沉默,还能在细胞分裂过程中稳定遗传,甚至在沉默诱导因子撤除后仍能维持。其优势在于单次转染即可实现高效率的靶基因沉默,并维持数十代细胞。然而,该系统对非CpG岛启动子效率较低,且分子量偏大。
CRISPRon作为其正交逆转系统,是利用dCas9-TET1融合蛋白催化DNA去甲基化,并整合MS2-p65-Rta辅助激活域实现协同激活。其中,TET1催化DNA的去甲基化,p65和Rta协同解除染色质抑制状态,sgRNA经改造插入MS2茎环,可招募MS2-p65-Rta融合蛋白。该系统可在72小时内逆转CRISPRoff诱导的沉默,使基因表达恢复至基线水平,且对高甲基化肿瘤抑制基因再激活效率达传统CRISPRa系统的3倍。
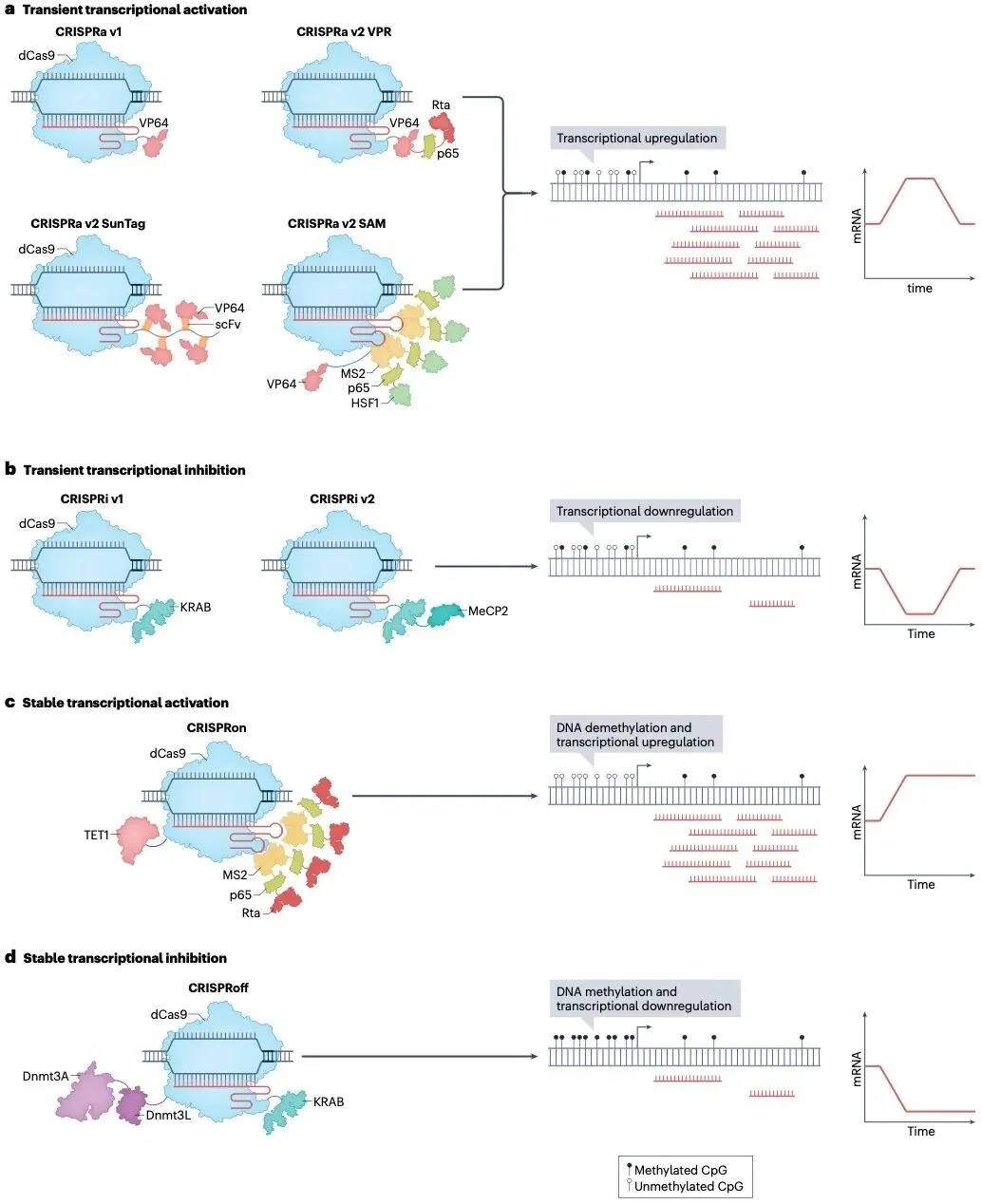
图4 | dCas9体系表观遗传编辑工具的转录调控
(来源:Nat Rev Mol Cell Biol. 2024;25(6):464-487.)
在ZFN和TALE基础上发展的表观遗传编辑工具虽不受PAM限制,但构建方法复杂。基于CRISPR-dCas9体系开发的工具极大地提高了靶向设计的便捷性,使表观遗传编辑技术得到了迅速发展。CRISPR体系表观遗传编辑工具的升级迭代体现了从单一效应域变为多效应域协同的趋势。早期基于dCas9体系的转录激活系统(如dCas9-VP64)在应用中面临显著的效能局限,其单效应结构域设计往往无法达到预期的基因激活水平,尤其在沉默染色质区域或低响应性启动子中表现更为显著。为突破此瓶颈,策略设计转向多重协同机制:一方面通过设计多靶点sgRNA同时靶向同一基因的启动子及远端调控元件(如增强子),利用空间邻近效应增强转录复合物的协同组装;另一方面则聚焦于dCas9蛋白的工程化改造,通过融合多价效应结构域(如VPR三联体)或构建模块化招募系统(如SAM、SunTag)实现效应分子的局部富集。此类第二代激活系统的核心创新在于效应功能的时空耦合,例如SAM系统通过sgRNA支架上的MS2茎环招募MS2-p65-HSF1复合物,与dCas9-VP64形成四元激活模块,在靶位点创造超高浓度的转录因子微环境,从而将激活效能大幅提升。然而,最优激活策略的选择呈现显著的环境依赖性,例如启动子类型(TATA盒依赖型/富含CpG岛型)、染色质基础状态(开放/封闭异染色质)、细胞类型(干细胞/终末分化细胞)及靶基因的天然表观记忆特性(如印记基因)共同决定了不同系统的适用性。
为了更适合于临床药物的开发,CRISPR体系表观遗传编辑工具的发展需要致力于能够提升其临床应用性的方向,例如开发紧凑型dCas变体,降低编辑工具的免疫原性,提高表观遗传编辑的效率和持久性,同时避免编辑工具在体内的长期暴露,规避脱靶效应,以及优化体内递送策略来实现临床转化。
三、表观遗传编辑技术的特点
基因组调控技术的多元化发展为精准医学提供了多层次干预策略。基因编辑技术(如CRISPR-Cas9)通过诱导DNA双链断裂(DSB)并依赖细胞内在修复机制,即非同源末端连接(NHEJ)或同源定向修复(HDR),实现对靶基因的永久性敲除或序列修正。其核心优势在于干预的彻底性和持久性,尤其在遗传性疾病(如镰状细胞病)的基因修正中展现临床潜力。然而,DSB引发的染色体异常风险(如大片段缺失、易位)及同源定向修复效率不足(<10%)构成主要安全性瓶颈。
碱基编辑技术在CRISPR框架下,通过脱氨酶融合的Cas9变体直接催化单碱基转换,规避DSB依赖的编辑方式,演进为无断裂修正工具,显著提升单碱基修正效率并避免了插入或缺失(indel)突变的产生。但碱基编辑存在编辑窗口限制(4~8 bp),当脱氨酶在靶向目标碱基时,可能对其编辑窗口内的非目标碱基进行非特异性修饰,即旁观编辑效应(bystander editing)。此外,碱基编辑工具的脱氨酶在无sgRNA引导的情况下,有可能随机结合基因组中瞬时暴露的单链DNA(ssDNA)区域并催化非目标位点的碱基转换,造成非序列依赖的脱靶编辑风险。
RNA干扰技术(RNAi)在转录后层面通过RNA诱导沉默复合物(RISC)降解目标mRNA,提供了一种快速且不会改变DNA序列的基因抑制方案。RNAi药物作用的可逆特性与体内递送方案的突破(脂质纳米颗粒(LNP)及GalNAc偶联)使其率先在临床应用中取得突破。自2018年Patisiran获批治疗转甲状腺素蛋白淀粉样变性以来,RNAi药物至今已走入成熟阶段,疾病领域也从遗传罕见病拓展到慢性病。然而,其药效作用时间虽然可持续数周至数月,相比传统药物形式已显著延长,但仍需反复给药,且易触发免疫应答(TLR激活)和脱靶基因沉默效应。
相比之下,表观遗传编辑技术是通过dCas9融合表观效应器定向重塑染色质状态,可在保持DNA序列不改变的前提下实现长效的基因调控。表观遗传编辑技术的核心优势在于其通过重塑染色质表观状态(如DNA甲基化、组蛋白修饰)实现基因表达的精准调控,完全规避DNA序列的物理改变。这一“无切割”特性可在保留基因组完整的状态下定向改写表观密码,从而避免因DSB引发的染色体易位、大片段缺失等不可控风险。值得注意的是,其安全性优势不仅体现于规避DSB相关的基因毒性,更在于脱靶效应的生物学影响显著低于基因编辑,因为表观遗传修饰的错误沉积通常可逆且极少干扰关键编码区功能。此外,该技术的可逆性也为治疗增加了一重安全机制。通过靶向同一基因座的反向效应器(如甲基化沉默后以去甲基化激活),可恢复基线表达水平,为剂量依赖毒性提供“紧急终止”机制。
在调控维度上,该技术展现出超越二元开关的高级可控性。通过设计差异化的效应器组合,可实现对基因表达水平的连续精细调控(剂量控制),解决传统基因编辑工具面临的“全有或全无”局限。例如血红蛋白病治疗中,通过甲基化强度梯度可精确提升胎儿血红蛋白(HbF)至25%-40%的治疗阈值,避免表达不足的情况下缺乏疗效或过表达后引发红细胞毒性的风险。
多重编辑能力是该技术的另一项关键优势。通过设计多靶向性的CRISPR系统(如阵列化gRNA),可同步重构多基因的表观状态,适用于多基因互作疾病,如自闭症谱系障碍或癌症表观协同突变。以Rett综合征为例,dCas9-TET1与dCas12-CTF联用成功激活MECP2并加固CTCF绝缘边界,实现双位点协同修复。该策略有效规避了基因编辑技术在多基因同步切割下引发的基因组不稳定性风险,为多基因遗传病治疗提供了新范式。
时空调控灵活性则依托表观记忆的可编程特性。通过选择可遗传性效应器(如CRISPRoff系统)可实现跨细胞分裂的持久沉默,而临时性效应器(如CRISPRon系统)支持瞬时激活后再恢复基线状态。这种“可遗传/可擦写”的双模式兼容临床需求多样性。例如在肿瘤免疫治疗中只需短暂激活检查点基因(如PD-1抑制剂响应期),或在遗传代谢病中永久沉默致病基因(如PCSK9终身沉默)。
尽管表观遗传编辑在靶向性、安全性和可调控性方面具有显著优势,其临床应用仍面临多重挑战。其作用位点的生物学约束构成首要限制。效应器虽可靶向启动子、增强子及绝缘子等调控元件,但编辑效率受局部染色质状态调控。异染色质区域因其致密结构导致靶向困难,而CpG岛虽为经典靶点,但仅占调控网络的有限部分。染色质标记在特定基因座上的线性扩散现象虽可扩大表观遗传调控作用范围,亦可导致距离靶位点数千个碱基对的超远端基因沉默,仍存在一定的风险。
靶点-功能映射的知识图谱尚不完整则是另一个瓶颈。多数的潜在调控位点目前还缺乏因果性验证,仅有少部分位点具备明确调控逻辑。表观遗传修饰的层级互作网络进一步增加调控复杂性。单一修饰可能受其他表观标记的反向调节,且组蛋白修饰与DNA甲基化间的动态串扰常导致编辑效果偏离预期。
编辑的持久性矛盾源于表观系统的固有动态平衡。甲基化修饰虽在启动子区相对稳定,但仍面临去甲基化酶的持续校正。细胞自主补偿机制也可能导致疗效削弱,如抑制DNA甲基化可能触发其他表观遗传修饰的代偿性上调,形成新型表观屏障。同时,表观遗传编辑器的环境依赖性决定了对不同位点存在效率差异。例如研究发现相同编辑器在SNURF位点可实现长期沉默,但同靶向策略对HER2仅短暂有效。这种差异源于染色质微环境(如三维构象、局部蛋白富集)对效应结合的空间阻遏效应。因此,在应用于不同的基因位点时,可能需要对靶向工具进行微调以获得最优效果。此外,表观遗传激活工具当前的效能偏低,基因激活效率普遍低于抑制,且依赖高乙酰化的染色质环境,尚不具备广泛应用的条件。
递送系统的适配性挑战直接制约体内应用。dCas9-效应器融合蛋白通常大于5 kb,超过了腺相关病毒(AAV)载体容量限制,转而需采用慢病毒或LNP的递送方式。
综上,各技术呈现边界清晰的特征谱:RNAi侧重瞬时干预的低风险性,碱基编辑聚焦点突变的精准修正,基因编辑瞄准根治性序列重构,而表观编辑则填补了“不改变基因、长效精准调控”的需求空白。未来技术迭代需在特异性、递送效率与时空控制三维度持续突破生物学边界。
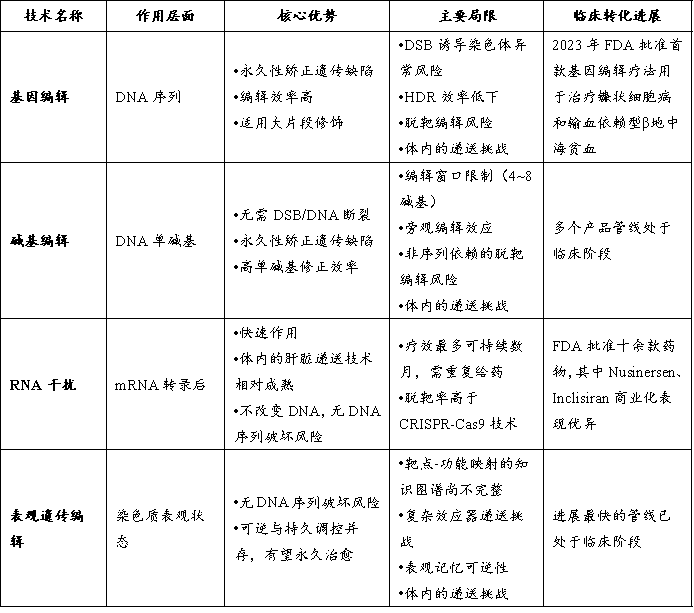
表2 | 各项技术对比
(来源:信熹资本整理)
四、表观遗传编辑的应用
表观遗传编辑技术已成为基础研究与转化医学的革新性工具,其在科研领域的应用深刻重塑了功能基因组学和细胞工程的范式。尽管已发现了数十种针对DNA和组蛋白的表观遗传化学修饰,但对于其中每种修饰的功能及其如何在基因组位置、细胞类型和环境条件下的变化通常仍所知甚少。表观遗传编辑技术在研究基因调控机制方面具有重要的应用价值。通过精确操纵特定基因位点的表观遗传标记,可建立表观修饰与基因功能的直接因果关联。典型案例如dCas9-DNMT3A融合蛋白在CTCF结合位点诱导DNA甲基化,直接证实了甲基化通过干扰CTCF结合破坏染色体拓扑结构域(TAD)边界的机制。在非编码基因组注释领域,基于CRISPRi和CRISPRa的高通量筛选已系统性解码人类细胞中数千个增强子的功能多样性,如Schmidt等发现24,000个候选增强子中的77%在特定CD8+T细胞亚群中具有特异性功能,揭示了基因调控的不同机制。
在细胞编程研究中,表观遗传编辑技术突破了传统转录因子重编程的局限性。Black等人利用CRISPR激活筛选技术,在小鼠胚胎干细胞(ESC)中鉴定出Ascl1、Dlx2等转录因子作为神经命运决定关键因子,通过dCas9-VP64靶向激活这些基因成功将成纤维细胞转分化为功能性神经元。在人类iPSC中的平行研究进一步揭示表观遗传修饰剂KDM6A(H3K27去甲基化酶)通过协同作用提升神经分化效率数倍,证实组蛋白修饰动态对谱系特化的调控作用。Yang等通过全基因组CRISPRa筛选发现TET1能将已分化的外胚层干细胞重编程为类多能态,其效率远超传统转录因子方法,且重编程细胞可稳定形成嵌合体胚胎,证明表观编辑策略具有跨谱系转化的普适性。体内转分化研究利用dCas9-VP16/p65系统直接将小鼠肝脏细胞重编程为胰岛素生成细胞,规避了传统方法中基因插入导致的致癌风险。
在免疫细胞工程领域,该技术通过表观重塑对抗或逆转T细胞耗竭以及改善细胞内在属性(如效应功能、增殖性和持久性),实现T细胞功能优化。dCas9-DNMT3A靶向抑制TET2基因可延缓CD8⁺T细胞耗竭表型,而dCas9-p300介导的IL-2增强子激活显著提升效应T细胞扩增能力。正交dSaCas9-p300系统可实现原代T细胞多重表观调控,筛选确定BATF-IRF4复合物为CD8⁺T细胞效应功能核心调控节点;体内实验证实靶向该通路的表观编辑可延缓T细胞耗竭并提升肿瘤清除率。
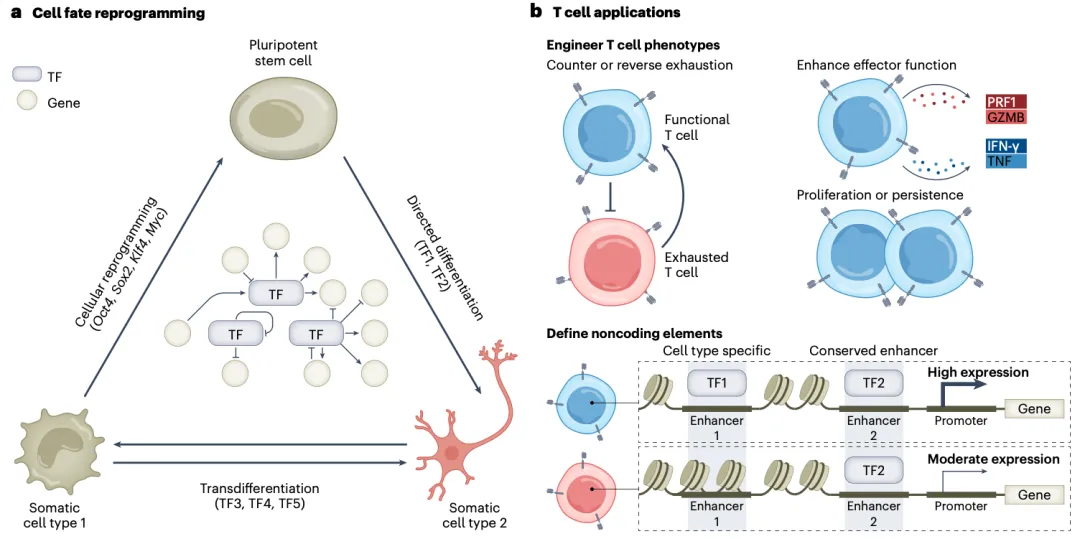
图5 | 表观基因组工程离体与在体应用
(来源:Nat Biotechnol. 2024;42(8):1199-1217.)
表观遗传编辑的独特优势在于其能解析复杂表观层级的动态互作。例如当靶向DNA去甲基化无法有效激活FMR1基因时,同步消除抑制性组蛋白标记可实现转录再激活,表明组蛋白修饰与DNA甲基化协同构成沉默屏障。
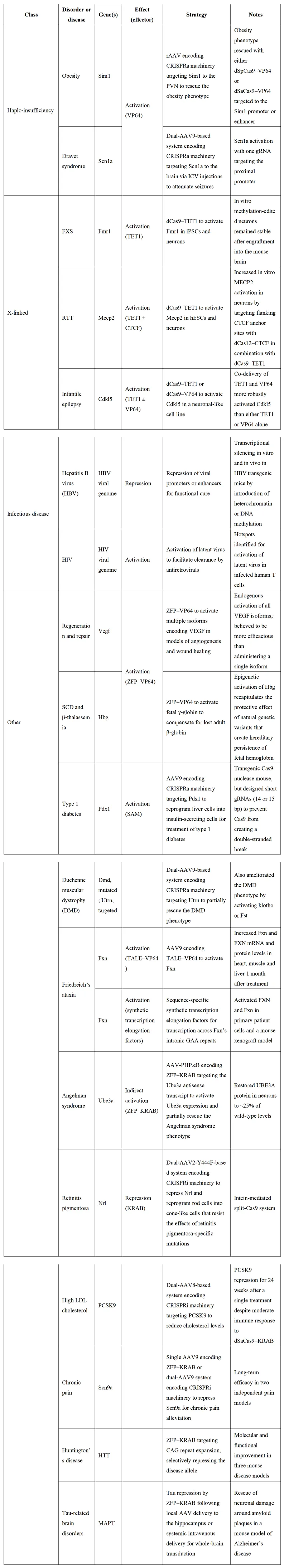
这些机制研究在针对源于多种表观遗传失调形式的疾病和促进再生医学方面具有巨大潜力,可促进疾病治疗新靶点、新机制的发现。同时,表观遗传编辑技术通过精准调控表观遗传修饰可实现病理基因表达逆转,在疾病治疗领域取得新的进展。在代谢性疾病领域,单次LNP递送靶向PCSK9的ZFP-KRAB-DNMT3A融合编辑器(EvoETR),可诱导小鼠肝脏持续基因沉默近1年,并维持低密度脂蛋白胆固醇(LDL-C)水平大幅降低。该效应在肝脏强制再生后仍稳定存在,证实表观修饰的遗传性。在非人灵长类研究中,人源PCSK9靶向表观编辑器单次给药实现持续1年的PCSK9蛋白水平降低90%,LDL-C下降70%,为高胆固醇血症提供革命性治疗方案。
针对神经发育疾病,dCas9-TET1介导的FMR1基因启动子去甲基化成功恢复脆性X综合征患者来源iPSCs的神经元功能,逆转CGG重复扩增导致的病理性基因沉默。在Rett综合征治疗中,dCas9-TET1靶向激活MECP2基因使野生型蛋白表达实现了一定程度恢复,显著改善神经元电生理异常。
在慢性病与感染性疾病领域,表观遗传编辑技术展现多维度疗效。在对糖尿病治疗的研究中,dCas9-VPR激活Pdx1基因使1型糖尿病小鼠肝细胞转分化胰岛素生成细胞。在疼痛领域,利用ZFP-KRAB靶向抑制Scn9a(钠通道基因)在小鼠疼痛模型中实现慢性疼痛缓解。在病毒感染治疗方面,表观遗传编辑技术为乙肝病毒(HBV)治疗提供了创新策略,其核心在于靶向沉默病毒共价闭合环状DNA(cccDNA)的表观遗传修饰,实现持久转录抑制。基于CRISPR-dCas9的系统通过融合KRAB或DNMT3A效应结构域,可特异性诱导HBV cccDNA的异染色质化或DNA超甲基化,从而抑制病毒复制。例如dCas9-KRAB靶向HBV CpG岛增强子区域,在HepG2.2.15细胞模型中将HBsAg表达降低90%以上(持续>6个月),并伴随H3K9me3修饰富集。这一策略在小鼠HBV转基因模型中验证了其体内有效性,单次给药后病毒载量下降约80%。Tune Therapeutics针对慢性乙型肝炎开发的疗法Tune-401目前已进入临床试验阶段。此外,VP64-p65的研究发现可重新激活潜伏的人类免疫缺陷病毒(HIV),以便通过抗逆转录病毒治疗清除病毒。
表观遗传编辑技术在其他疾病方面也做了积极探索。例如ZFP-VP64激活胎儿γ-球蛋白补偿β-地中海贫血患者成人血红蛋白缺陷,有望使输血依赖患者长期摆脱输血。TALE-VP64恢复Friedreich共济失调患者细胞中frataxin蛋白表达,改善心肌线粒体功能。ZFP-KRAB靶向UBE3A-AS反义RNA使Angelman综合征模型神经元UBE3A蛋白恢复至野生型25%,在Angelman综合症小鼠模型中部分恢复了探索性运动能力。
表3 | 表观基因组编辑技术在人类疾病治疗中的应用
(来源:Nat Biotechnol. 2024;42(8):1199-1217.)
五、表观遗传编辑的挑战
表观遗传编辑技术当前面临的核心挑战涵盖机制认知、技术限制及临床应用三方面。表观遗传编辑技术的发展受限于靶点-功能映射知识图谱的显著缺口。当前研究仅验证了约12%的疾病相关表观遗传位点的直接调控功能,而高达88%的潜在调控位点(如增强子、沉默子)尚未建立明确因果逻辑。例如前列腺癌中77%的AR(雄激素受体)相关增强子仅通过染色质共定位预测功能,但dCas9-KRAB靶向筛选显示仅35%可实际调控AR转录。这种认知缺失导致编辑策略设计缺乏高效的科学规则。表观遗传修饰机制解耦难题进一步复杂化靶点验证,构建表观遗传修饰与转录变化的关联性仍然是一个重要的研究领域。相关研究发现,dCas9-p300在HEK293T模型中诱导H3K27ac富集仅能使60%-70%的靶基因激活,且H3K4me1写入增强子后仅40%触发预期染色质环重构。混合筛选发现VPR激活结构域虽在部分基因位点提升效率数倍,但难以推广至全基因组。因此,明确表观修饰与基因表达的直接调控关系是进一步开发利用该技术的前提。
细胞环境依赖性显著影响编辑效率。CpG岛密度差异导致编辑效率显著不同,在CpG贫乏区域(如HBV cccDNA)的甲基化编辑效率可能仅为CpG富含区域的20%。细胞类型特异性的表观遗传标记可以影响dCas9-DNA结合动态,使得效果在不同细胞类型中不一致。此外,研究发现神经发育关键期(如出生后4周内)的染色质可及性变化使Syt1增强子编辑效率从新生小鼠上的82%骤降至成年小鼠上的11%。在实体瘤微环境中,缺氧诱导的HIF-1α显著削弱dCas9-DNMT3A对GSTP1的再甲基化能力。
递送系统是影响表观遗传编辑应用于的产品开发的关键因素。AAV载体存在容量限制(容纳序列<4.7kb),而目前开发的表观遗传编辑工具的结构都较复杂,其体积超过AAV可容纳的上限,并且病毒载体激发的中和抗体为重复给药制造了障碍。LNP递送虽可承载全长蛋白,但进入体内后易富集于肝脏,非肝组织的分布率低,影响了肝外靶点的编辑效率。
长效性与剂量控制问题并存。尽管靶向PCSK9的ZFP-KRAB-DNMT3A在小鼠中维持沉默超过1年,但脆性X综合征模型的FMR1去甲基化仅持续2周。对于基因表达水平敏感性疾病,基因激活或抑制水平的精确控制就显得非常重要。对于一部分基因来说,超生理表达可能不是问题,但对于某些基因则需要精确的剂量控制,因为在单倍体不足和过量的情况下都可能引发生理异常,例如MECP2过表达会诱发癫痫。相反,基因激活或抑制的不足则可能无法产生治疗效果。
针对这些挑战,研究者正在探索相应的解决方案来推动从技术端向临床端的转化。例如在递送方案优化方面,选择工程化小型化Cas蛋白(如dSaCas9)来缩小表观遗传编辑工具的体积,以及开发可靶向肝外组织的LNP递送系统。这些进展标志着表观编辑从机制研究迈向精准医疗的新阶段。
六、表观遗传编辑的代表企业
1、 Chroma Medicine(nChroma Bio)
Chroma Medicine是一家位于美国马萨诸塞州剑桥的生物技术公司,成立于2021年。该公司专注于表观遗传编辑领域,致力于开发新型基因组药物,通过调控基因表达来治疗基因驱动疾病,而不改变DNA序列本身。其核心技术是模块化表观遗传编辑平台,该平台将DNA结合域与表观遗传效应结构域结合,可精确编程以实现靶向基因的沉默、激活或多基因同时调控。
公司由多位基因组研究领域的顶尖科学家联合创办,包括David Liu(刘如谦)、Luke Gilbert、Keith Joung、Jonathan Weissman、Luigi Naldini和Angelo Lombardo。其中,David Liu是CRISPR先驱,碱基编辑和先导编辑技术发明者;Luke Gilbert是表观遗传调控机制专家;Jonathan Weissman是基因表达调控领域的专家。
2024年12月,公司与Nvelop Therapeutics合并成立nChroma Bio,整合表观遗传编辑与下一代体内递送技术,拓展肝外疾病治疗。
1)研发技术平台
Chroma Medicine的核心技术基于表观遗传编辑,通过可编程编辑器在不改变DNA序列的情况下调控基因表达。其平台的关键特点包括:
模块化编辑器设计。结合DNA结合结构域与表观遗传效应结构域,精准靶向基因并调控其活性。例如甲基化酶或去甲基化酶可分别用于基因沉默或激活,模仿细胞天然的基因调控机制。
避免DNA切割。与传统CRISPR技术不同,Chroma Medicine的编辑器通过表观遗传修饰(如甲基化)控制基因表达,减少脱靶效应和免疫原性风险。这一特性使其在安全性和监管合规性上更具优势。
多基因调控能力。平台支持同时靶向多个基因,适用于复杂疾病(如癌症、代谢性疾病)的治疗。
优化的TET分子库。通过迭代优化,Chroma Medicine开发了高效的去甲基化酶TET变体,其效力比已发表版本提升约4倍,可实现基因的持续激活。
2)研发管线进展
与Nvelop Therapeutics合并成立nChroma Bio后,首推项目为CRMA-1001,一种肝脏靶向表观遗传编辑疗法,目标为慢性乙型肝炎(HBV)和丁型肝炎(HDV)的功能性治愈。临床前数据显示其对HBV标志物的持久沉默效果显著。公司已启动首个临床候选药物CRMA-1001,标志着从平台技术向疾病疗法的转化。

图6 | nChroma Bio公司管线
(来源:公司官网)
3)融资情况
2021年11月:A轮融资1.25亿美元,由Cormorant Asset Management领投,用于平台开发和早期管线。
2023年3月:B轮融资1.35亿美元,GV领投,推动临床前研究向临床转化。
2024年12月:合并后融资,nChroma Bio获得7500万美元,支持CRMA-1001及肝外管线开发。
2、 Tune Therapeutics
Tune Therapeutics成立于2021年,由基因编辑领域的科学家Fyodor Urnov、基因调控专家Charles A. Gersbach以及生物技术运营与融资专家Akira Matsuno共同创立。公司总部设在美国西雅图,并在北卡罗来纳州达勒姆设有研发中心。公司主要聚焦表观基因组编辑(epi‑editing),致力于开发针对慢性疾病的基因调控疗法,尤其是慢性乙型肝炎等常见、慢性及老龄化相关疾病。
1)研发技术平台
Tune Therapeutics的核心技术为TEMPO(基因调谐平台)表观遗传编辑平台,该平台通过调节基因表达(上调或抑制)来治疗疾病,无需切割或永久改变DNA序列。TEMPO采用LNP递送系统,可同时靶向多个基因,适用于从罕见病到慢性病(如癌症、乙型肝炎、衰老相关疾病)的广泛领域。其独特之处在于精准调控表观基因组,逆转细胞功能异常,例如在非人灵长类动物中实现PCSK9基因的持久抑制,显著降低LDL胆固醇水平。
2)研发管线进展
Tune-401:针对慢性乙型肝炎的临床阶段表观遗传沉默药物,通过靶向病毒DNA(intDNA和cccDNA)抑制病毒复制。2024年11月获准在新西兰和中国香港开展临床试验。
PCSK9基因项目:2023年ASGCT会议上公布数据,单次治疗后非人灵长类动物的LDL胆固醇水平持续降低近两年,展示了表观编辑在心血管疾病中的潜力。
其他管线:包括基因、细胞和再生疗法项目,具体靶点尚未完全披露,但公司计划扩展至更多慢性疾病领域。
3)融资情况
2021年12月:A轮融资4000万美元,由New Enterprise Associates、Emerson Collective领投,用于TEMPO平台开发和临床前研究。
2025年1月:B轮融资1.75亿美元,由New Enterprise Associates、Yosemite、Regeneron Ventures和Hevolution Foundation领投,资金用于推进Tune-401临床试验、扩展管线及平台技术。
3、 Omega Therapeutics
Omega Therapeutics是一家总部位于美国马萨诸塞州剑桥的临床阶段生物技术公司,成立于 2016 年,由Flagship Pioneering孵化。公司核心使命是通过其核心技术平台,利用可编程的表观遗传药物在转录前精准调控基因表达,从而纠正疾病根本原因而不改变原生核酸序列。
1)研发技术平台
Omega Therapeutics的核心技术为Omega Epigenomic Programming™平台,通过设计模块化的mRNA编码表观遗传控制器(OEC),在不改变DNA序列的前提下精准调控基因表达。该平台利用LNP递送系统,将包含DNA结合域和表观效应器的mRNA递送至细胞内,针对传统疗法难以触及的靶点(如MYC致癌基因)进行干预。其技术优势在于可编程性,能够覆盖98%的“黑暗基因组”(非编码区域),为癌症、代谢性疾病等提供新疗法。
2)研发管线进展
Omega Therapeutics的研发管线包含8个临床前项目,覆盖以下领域:
肿瘤学:核心候选药物OTX-2002针对肝细胞癌(HCC)的MYC基因,在临床前模型中可显著抑制MYC表达,已进入临床1期。
代谢性疾病:与诺和诺德合作开发表观遗传控制器,通过增强能量消耗(如产热)治疗肥胖症,预计进入临床前开发。
其他领域:包括再生医学、炎症性疾病(如ARDS)、脱发、非小细胞肺癌等,计划每年新增2-3个候选药物。
3)对外合作
2023年,与诺和诺德达成价值高达5.32亿美元(包括前期、开发和商业里程碑付款)的合作,利用Omega平台开发肥胖管理疗法,诺和诺德承担研发费用并拥有独家商业化权利。
4)融资情况
2020年7月:A轮融资获Flagship独家8500万美元。
2021年3月:C轮融资1.26亿美元,由Flagship领投,BlackRock、Fidelity等跟投。
IPO进展:2021年7月登陆纳斯达克,代码OMGA,总融资额达2.11亿美元。公司现已退市。
4、 Epic Bio
Epic Bio成立于2018年,总部位于美国加利福尼亚州南旧金山,由亓磊(Stanley Qi)教授和Amber Salzman博士共同创立。公司致力于通过其核心平台Gene Expression Modulation System(GEMS)——一种基于极小型Cas蛋白CasMINI的非切割式基因调控技术,实现单一AAV载体递送,以精准激活或抑制基因表达,填补目前缺乏有效疗法的疾病空白。
1)研发技术平台
Epic Bio的核心技术为基因表达调控系统(GEMS),可通过体内或体外(如CAR-T细胞疗法)应用,动态调控基因表达,无需永久改变DNA。该平台结合三大组件:
调节剂:包括激活剂、阻遏剂及表观遗传调节剂(如组蛋白修饰剂、DNA甲基化/去甲基化剂),形成庞大的调节剂库,可精准控制基因表达剂量并引入持续变化。
DNA结合蛋白:采用从斯坦福大学授权的超紧凑CasMINI蛋白,尺寸仅为Cas9/Cas12a的一半,可通过单链AAV载体递送至多种器官,降低免疫原性并提高特异性。
引导RNA:通过功能筛选平台设计高特异性引导RNA,最大限度减少脱靶效应。
2)研发管线进展
公司目前推进5条临床前管线,覆盖多种遗传疾病:
EPI-321:针对面肩肱型肌营养不良症(FSHD),获得FDA“孤儿药”资格,已进入临床1期。
其他适应症:杂合子家族性高胆固醇血症(HeFH)、α-1抗胰蛋白酶缺乏症(A1AD)、色素性视网膜炎4型(RP4)及11型(RP11)。
此外,Epic Bio与吉利德子公司Kite Pharma合作开发下一代CAR-T疗法,旨在增强持久性与疗效。
3)融资情况
2022年7月:A轮融资5500万美元,由Horizons Ventures领投,用于推进临床前管线及技术开发。
2023年4月:金额未披露,投资方包括SOLVE FSHD。
2025年3月:B轮融资6800万美元,由汇桥资本领投,支持FSHD等适应症研发。
七、总结
表观遗传编辑技术作为一种新兴的精准调控工具,在基因治疗领域展现出独特的潜力和挑战。与成熟的基因编辑(如CRISPR-Cas9)、碱基编辑或RNAi技术相比,其技术成熟度仍处于发展初期,但核心优势在于能够在不改变DNA序列的前提下,实现对基因表达的长期甚至永久性调控,为单次治疗提供持久疗效。尽管脱靶效应不可避免,但优化后的表观编辑器脱靶率显著低于传统基因编辑工具,且因表观修饰具有可逆性,轻微脱靶通常可由细胞自身机制修复,总体安全性较高。该技术适合开发体内疗法,尤其适用于表观遗传失调相关疾病(如癌症、神经退行性疾病)及慢性病管理,因其通过调控基因表达而非修正突变,在体内治疗中兼具长效性和安全性优势;然而,对于基因突变的功能缺失型(LOF)遗传病,基因编辑或碱基编辑通常仍更具针对性。在体外应用方面,该技术目前则更多局限于基础机制研究。
表观遗传编辑药物开发面临靶点选择限制,受到CpG岛表观敏感区域、表观修饰与基因表达的直接调控相关性,以及不同修饰间互作网络的影响。因此,当前早期阶段适合以机制明确的靶点(如PCSK9用于血脂调控、乙肝病毒沉默)以降低开发风险。值得注意的是,RNAi技术已实现长效给药(如数月间隔),因此表观编辑药物需在疗效持久性或显著性上超越现有方案才具临床竞争力。递送系统是当前主要瓶颈,LNP因其在mRNA及siRNA疗法中的成熟应用成为首选载体,但肝外组织的特异性递送仍需等待突破,目前业界在靶向T细胞、肺部等组织器官方面已出现进展;病毒样颗粒(VLP)等新兴技术虽具潜力,但表观编辑器的封装和表达稳定性仍是挑战。总体而言,表观遗传编辑技术有望通过递送创新与靶点优化,推动下一代基因疗法的临床转化。
参考资料:
Nat Biotechnol. 2024;42(8):1199-1217.
Nat Cell Biol. 2021;23(1):11-22.
Epigenetics. 2022;17(5):564-588.
Nat Rev Mol Cell Biol. 2024;25(6):464-487.
Curr Opin Chem Biol. 2020;57:75-81.
END
延伸阅读
信熹洞见 · 研究
信熹对话
谱域科技创始人解安亮:以终端破局,打通卫星互联网的“最后一公里”
信熹伙伴
北外滩科创会客厅
